Вплив генотипу на потенціал зберігання цукрових буряків (частина 2)

За матеріалами наукової статті на тему «Інтегративна транскриптоміка виявляє генотипний вплив на придатність цукрових буряків до зберігання» («Integrative transcriptomics reveals genotypic impact on sugar beet storability») міжнародного журналу з молекулярної біології, молекулярної генетики та біохімії «Молекулярна біологія рослин» («Plant Molecular Biology»), серпень 2020 р.
Зміни під час зберігання з урахуванням потенціалу зберігання
Для ідентифікації DEGs між добре і погано придатними до зберігання сортами для кожної часової точки було обчислено лінію перетину DEGs між усіма добре та погано придатними до зберігання сортами (V1 vs V2, V1 vs V5, V6 vs V2 та V6 vs V5), в результаті чого було отримано 82 DEGs під час T0, меншу кількість під час Т1 та Т2 (30 та 36 DEGs відповідно) та до 905 DEGs під час Т4 (Рис. 5а, b). Всього було виявлено 1 011 диференційовано експресованих генів між добре та погано придатними до зберігання сортами. Взагалі, гени сортів, які погано зберігаються, частіше виявляються вищими, ніж у сортів, які добре зберігаються, та навпаки. Серед 1 011 генів було визначено лише шість генів, які вважалися диференційовано експресованими між добре і погано придатними до зберігання сортами у всі часові точки, два з яких мають функціональну анотацію, білок F-box у 5 u 03970-подібному гені та білок-транспортер органічних аніонів/карнітину 2 з гомологом арабідопсису OCT5.
Рис. 5. Порівняння хороших та погано придатних до зберігання сортів. Діаграма Венна, що показує кількість диференційовано експресованих генів між добре і погано придатними до зберігання сортами в кожен момент часу (а). Гістограма кількості DEGs у кожен часовий момент (b). Аналіз генної онтології (GO) негативно (c) та позитивно регульованих генів (d) під час Т4 відповідно до біологічних процесів, візуалізованих за допомогою REVIGO.
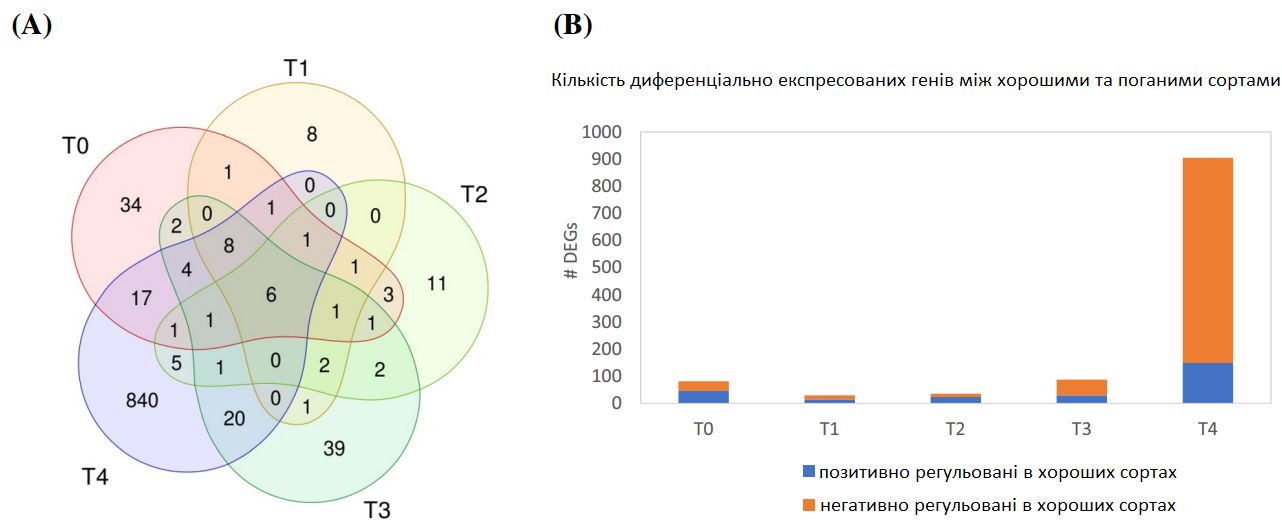

Зосереджуючись на Т0 (збирання врожаю), до початку зберігання можна було виявити 82 гени, розрізняючи добре та погано придатні до зберігання сорти. Була виявлена позитивна регуляція двох генів, відповідальних за поділ клітин (білок TORNADO 2, білок кінетохор гомолог NUF2), та два негативно регульовані гени, що беруть участь у рості клітин (Proflin-1, білок сімейства рамногалактуронат ліази) у добре придатних до зберігання сортах, порівняно з сортами, які погано зберігалися. Крім того, було виявлено, що гени беруть участь в реакції на абіотичні стреси, включаючи модифікації клітинної стінки, наприклад дві глікозилтрансферази, глутатіон-S-трансферазу та передбачувану бета-глюкозидазу 41, але також пов'язані із захистом проти грибів, наприклад AX1 та LYM2, які були інтенсивніше вираженими в сортах, які погано зберігалися.
На відміну від Т0, після 13 тижнів зберігання (Т4) було виявлено 905 диференційовано експресованих генів між добре і погано придатними до зберігання сортами (Рис. 5). Добре придатні до зберігання сорти включали гени, які належать до біологічних процесів метаболізму стеринів, біосинтезу глюкану та гіперосмотичної реакції клітин (Рис. 5). З іншого боку, погано придатні до зберігання сорти містили гени, які належать до процесів окислення-відновлення, включаючи клітинне розпізнавання та метаболізм вуглеводів, а також регулювання реакції на біотичний стимул для біологічних процесів (Рис. 5) та компоненти клітинної стінки рослинного типу, мембрани та позаклітинну ділянку, а також мітохондріальний протонотранспортний комплекс АТФ-синтази.
Серед топ-20 позитивно регульованих DEGs у добре придатних до зберігання сортах під час Т4 серед генів без анотації було виявлено два гени, які беруть участь в метаболізмі крохмалю (крохмаль-синтаза 1, альфа-глюкан-фосфорилаза), та гени, важливі для оксидативного стресу та реакції на патогенів, наприклад ймовірна тимет олігопептидаза, метиламіноксидаза міді, а також гени, що позитивно регулюють проліферацію клітин (гомолог арабідопсису в 3 г 07870; ELP4), та один ген, що бере участь у біосинтезі стеринів (ймовірний 3-бета-гідроксистероїд-дельта (8), дельта (7)-ізомераза). Гени, важливі для біосинтезу фенілпропаноїдів або фавоноїдів, були дуже низько регульованими, наприклад фенілаланін аміак-ліаза, 3-О-метилтрансфераза кавової кислоти, метилентетрагідрофолат редуктаза 1, гени ремоделювання клітинної стінки (основний 7S глобулін; альфа-глюкозидаза; та гени реакції на стрес (глутатіон S-трансфераза; білок резистентності до плейотропного препарату 1). Розглядаючи всі позитивно регульовані DEGs у добре придатних до зберігання сортах під час Т4, було виявлено набагато більше генів, пов'язаних із стресом і захистом рослин, включаючи шість (ендо) генів хітинази та дев'ять факторів транскрипції WRKY, деякі з них регулювали реакцію на стрес (WRKY6, WRKY15), а інші відповідали за захист рослин від патогенних мікроорганізмів (WRKY33, WRKY40, WRKY46). Крім того, було виявлено 13 герміноподібних білків (GLP), які, як відомо, мають функцію оксалатоксидази в цукрових буряках, що призводить до утворення пероксидів водню, другої сигнальної молекули під час реакції рослин на стрес, та шість білків DIR, які, як вважається, можуть утворюватися під дією стресу та брати участь у контролі метаболізму клітинної стінки та/або формуванні антибактеріальних сполук.
Згідно з DEGs, пов’язаними з процесом дихання, було виявлено вісім негативно регульованих генів у добре придатних до зберігання сортах після 13 тижнів зберігання (Т4) у шляху гліколізу: гексокіназа-1, фруктоза та глюкозо фосфорилюючий фермент, який регулює ріст і розвиток рослин незалежно від цукрового обміну, та АТФ-залежну 6-фосфофруктокіназу, яка, як відомо, відіграє важливу роль у гліколізі цукрових буряків після збирання врожаю. Було нараховано десять диференційовано експресованих генів у крохмалі та сахарозі, включаючи три позитивно регульовані гени, пов'язані з крохмаль-синтазою у хороших сортах. У добре придатних до зберігання сортах також було виявлено регуляцію двох генів цитратного циклу (дигідроліпоїл дегідрогенази, мітохондріального та АТФ-цитрат-синтази білка альфа-ланцюга 1) та три гени в ланцюжку транспорту електронів та комплексу АТФ-синтази (протохема IX фарнезилтрансфераза, АТФ-синтаза субодиниця дельта та епсилон), що підтверджує більше виробництво енергії у погано придатних до зберігання сортах після 13 тижнів зберігання.
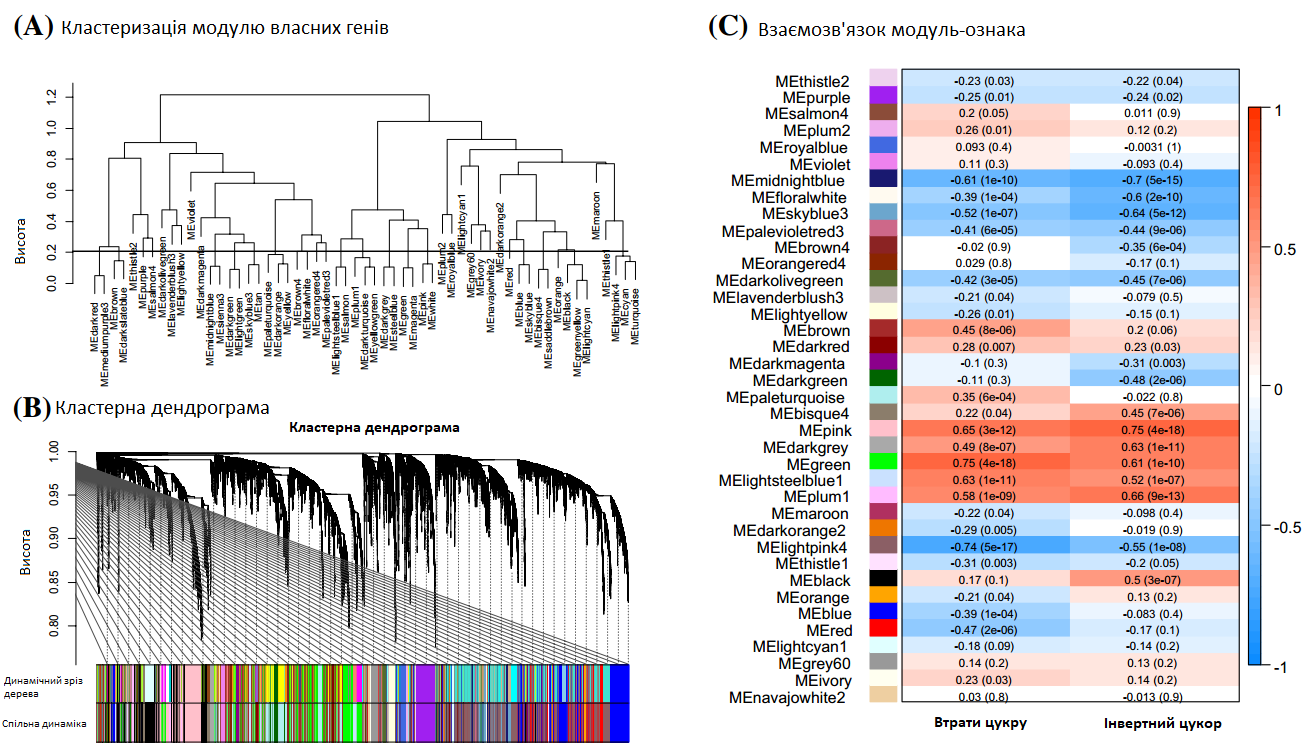
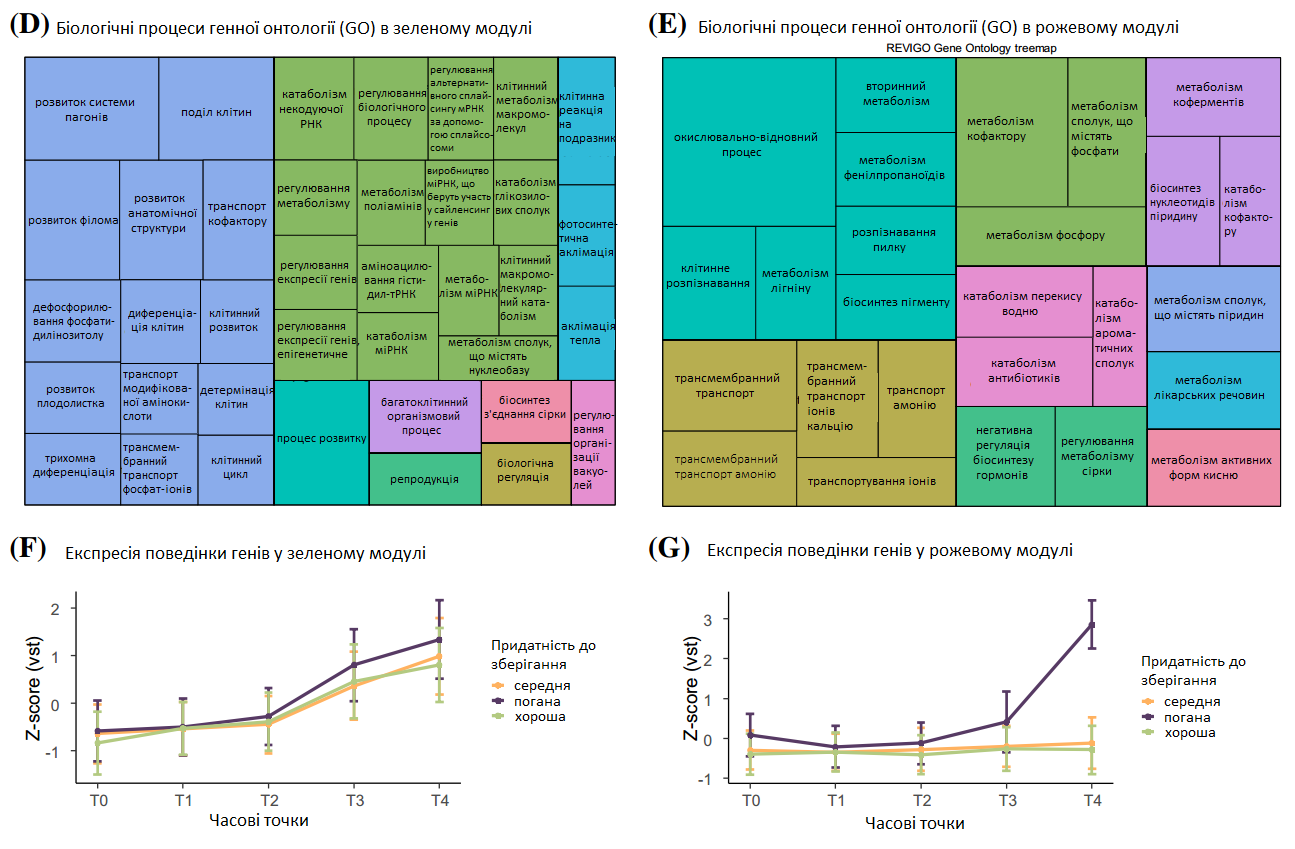
Рис. 6. Аналіз зважених мереж експресії генів (WGCNA). Кластерні модулі власних генів, ідентифіковані WGCNA та вирізані для об’єднаних модулів (а). Ієрархічне кластерне генетичне дерево, що показує модулі коекспресії (b). Основні гілки дерев утворюють 38 об’єднаних модулів, які позначені різними кольорами. Теплова карта, де кожен колір клітини показує кореляцію ознаки з кожним модулем власних генів WGCNA (c). Аналіз генної онтології (GO) у зеленому (d) та рожевому модулях (e) згідно біологічних процесів, візуалізованих за допомогою REVIGO. Профіль експресії всіх генів зеленого модуля корелює з втратою цукру (f), а рожевого модуля — з інвертним цукром (g). Було визначено медіану дисперсійних стабілізуючих значень трансформованої експресії для всіх зразків, що належать до однієї групи зберігання кожного гена та обчислено z-бал. Графіки показують середні значення z-балів усіх генів, а планки похибок показують стандартне відхилення.
Аналіз шляху KEGG
Аналіз KEGG значущих диференційовано експресованих генів між добре і погано придатними до зберігання сортами під час Т4 показав, серед іншого, збагачену негативну регуляцію хороших сортів у біосинтезі вторинних метаболітів, включаючи біосинтез фавоноїдів та фенілпропаноїдів, що призводить до біосинтезу лігніну, а також у сигнальному шляху мітоген-активованої протеїнкинази. Позитивна регуляція у добре придатних до зберігання сортах спостерігається в метаболічних шляхах загалом, а особливо в метаболізмі тіаміну. Цікаво, що в хороших сортах всі стадії шляху фенілпропаноїдів були негативно регульованими (мається на увазі позитивне регулювання у поганих сортах) виключно під час Т4 (Рис. 5).
WGCNA
Було проведено аналіз WGCNA, що включав усі зразки, які також використовувалися для DEG-аналізу. 19 551 генів було згруповано у 26 об’єднаних модулів (Рис. 6а, b), а модулі з високою кореляцією до найважливіших фізіологічних параметрів, що визначають придатність до зберігання, інвертний цукор та втрати цукру, були додатково проаналізовані (Рис. 6c). Найвища позитивна кореляція втрат цукру під час зберігання була виявлена в зеленому модулі (1 884 гени, r = 0,75, p = 4e – 18) з клітинним поділом та розвитком відповідних генів (Рис. 6d), а висока негативна кореляція була в рожевому модулі (3 140 генів, r = - 0,74, p = 5e – 17) з регуляцією захисної реакції на бактерії (Рис. 6a). Найвища кореляція інвертного цукру спостерігалася в рожевому модулі (1 086 генів, r = 0,75, p = 4e – 18), що складається з генів, які демонструють вторинний метаболізм (лігніну, фенілпропаноїдів), метаболізм коферментів або метаболічні процеси активних форм кисню (Рис. 6e). У свою чергу негативна кореляція (436 генів, r = -0,7, p = 5e - 15) була надана темно-синьому модулю, який демонструє, серед іншого, клітинну реакцію на абіотичний стимул та процес метаболізму азотних сполук. Гени в зеленому модулі демонструють загальне збільшення експресії протягом періоду зберігання (Рис. 6f), тоді як гени в рожевому модулі демонструють значне збільшення експресії під час Т4 виключно у поганих сортах (Рис. 6g). Далі визначалися гени-концентратори висококорельованих генів, беручи до уваги внутрішньомодульний зв’язок аналізу WGCNA та додатково ступінь зв’язку відомих взаємодій білок-білок. Можна визначити п’ять генів-концентраторів, які пов’язані з втратою цукру, і шість генів, пов’язаних з інвертним цукром. Гени-концентратори втрати цукру включають три гени, які діють як шаперони, так і з ними (білок dnaJ ERDJ2A, фактор обміну нуклеотидів SIL1, фактор елонгації транскрипції гомолог SPT6), та ген, що бере участь у гліколізі (триосефосфат-ізомерази, цитоплазми). Гени інвертного цукру включають три синтази халкону та катехол-CoA O-метилтрансферазу.
Обговорення
У даному дослідженні було проведено порівняльну транскриптоміку цукру та стандартного аналіту, а також анатомічні аналізи коренеплодів шести сортів цукрових буряків під час зберігання. Було виявлено фактори та механізми з різноманітним впливом на придатність до зберігання, які змогли розрізнити добре і погано придатні до зберігання сорти та їх характер протягом 13 тижнів зберігання.
Використання цукру для боротьби зі стресом від посухи та нападами патогенів — загальний ефект під час зберігання
Як правило, протягом усього періоду зберігання у всіх сортах буряків вміст цукру (SC) зменшувався, тоді як кількість інвертного цукру збільшувалася, що є основними проблемами цукрової галузі. Крім того, під час зберігання було виявлено загальне збільшення кількості клітин паренхіми, що підтверджувалося експресією генів, важливих для поділу клітин, тоді як відбувалося перешкоджання росту клітин (негативне регулювання семи ксилоглюканових ендотрансглюкозилаз) та зменшення цілісності клітинної стінки, ймовірно, через негативне регулювання семи лакказ. Відомо, що лаккази беруть участь у біосинтезі та деградації лігніну, але після зберігання не було виявлено суттєвих відмінностей у загальному вмісті лігніну. Таким чином, найбільший рівень експресії лакказ під час Т0 також можна пояснити реакцією загоєння ран, яка відбулася безпосередньо після збирання врожаю. Крім того, під час зберігання було виявлено загальне збільшення товщини перидерми, швидше за все, для більш контрольованого регулювання обміну води та газу, а також для поліпшеного захисту від біотичних та абіотичних стресів, особливо після механічних пошкоджень коренеплодів під час збирання. Це підтверджується високою позитивною регуляцією генів, які необхідні для збільшення стійкості до осмотичного та посухового стресу, для боротьби з водним стресом під час зберігання. Оскільки перидерма складається з пробкових клітин, очікувалося експресування важливих ферментів у фенілпропаноїдному шляху під час зберігання. Однак дане дослідження підтвердило попередні висновки, що синтез перидерми в цукрових буряках не пов'язаний з активністю фенілаланін-аміак-ліази (PAL) та пероксидази (POD).
Здається, що загальні втрати цукру під час зберігання в цукрових буряках поєднуються зі зміною стратегії росту, яка характеризується зменшенням розширення клітин та збільшенням їх поділу з подальшим розвитком перидерми. Вже було доведено, що під час зберігання завжди відбувається фізіологічна та молекулярна реакція на абіотичний стрес. Ми також виявили підвищену експресію генів, пов'язаних із захистом від патогенів під час зберігання, однак стратегію захисного реагування, загальну для всіх шести досліджуваних сортів, побачити не вдалося, що, ймовірно, вказує на специфічні для кожного сорту механізми реагування.
Передумови для добре придатного до зберігання сорту
Сорти, які краще зберігаються, мали більший вміст цукру, більшу кількість камбіальних кілець та клітин паренхіми, тоді як розмір самих клітин був меншим, у порівнянні з погано придатними до зберігання сортами, під час Т0. Це можна підтвердити порівняльною транскриптомікою, де два позитивно регульовані гени, що відповідають за поділ клітин, а також два негативно регульовані гени, що беруть участь у рості клітин, були виявлені у добре придатних до зберігання сортах.
Добре придатні до зберігання сорти мали тоншу перидерму разом із меншим вмістом лігніну. Оскільки лігнін, як відомо, є важливим бар'єром для патогенних мікроорганізмів, можна припустити, що погано придатні до зберігання сорти мали потребу у встановленні товстіших клітинних стінок раніше в процесі свого розвитку, щоб зменшити чутливість до нападів патогенів, що може бути викликано значним підвищенням регуляції DIR23, гену, що бере участь у біосинтезі лігніну, який негативно регулюється у добре придатних до зберігання сортах. Крім того, хороші сорти, здається, оснащені іншою, очевидно, більш ефективною захисною системою, що, ймовірно, викликано значним позитивним регулюванням члена сімейства генів бета-глюкозидази (BGLU41), що вказує на негайний хімічний захист від патогенів. Крім того, було виявлено, що більший вміст поживних речовин пов’язаний з хорошою придатністю до зберігання, що доводить твердження про те, що вищий вміст поживних речовин спостерігався ще до початку зберігання.
Підводячи підсумок, можна сказати, що під час збирання врожаю (Т0), перед зберіганням, добре і погано придатні до зберігання сорти можуть мати різну реакцію на абіотичні стреси, серед іншого, демонструючи різну експресію генів модифікації клітинної стінки, а також різну реакцію на напади патогенів.
Погана придатність до зберігання поєднується з підвищеною реакцією на стрес та захистом від патогенних мікроорганізмів
Під час зберігання позитивне регулювання метаболізму крохмалю у добре придатних до зберігання сортах видається дуже цікавим, оскільки відомо, що цукровий буряк зберігає енергію лише у формі цукру, а не крохмалю, незважаючи на вираження генів біосинтезу крохмалю. Однак геном, який виявився позитивно регульованим у хороших сортах, була альфа-глюкан-фосфорилаза, яка є частиною процесу метаболізму крохмалю, який, як відомо, витримує дефіцит водного стресу, але не змінює вміст крохмалю в арабідопсисі. Хороші сорти краще справляються зі станом гіперосмотичних клітин, швидше за все, через водне напруження під час зберігання, а також збільшення реакції на оксидативний стрес. Цікаво, що мітохондріальна дигідроліпоїлдегідрогеназа була негативно регульованою у добре придатних до зберігання сортах, що свідчить про жорстку регуляцію клітинного дихання під час зберігання між хорошими та поганими сортами.
Наступним фактором, який може значно впливати на придатність сорту до зберігання, є вміст альфа-аміно азоту — чим вищий, тим краще для гарного зберігання. Відомо, що рослини здатні швидко реагувати на стресові фактори, збільшуючи концентрацію сумісних розчинених речовин, що беруть участь в осморегуляції, таких як азотні сполуки. Цілком ймовірно, що хороші сорти накопичують альфа-аміно азот для захисту коренеплодів від осмотичного стресу, що виникає під час зберігання. Однак значне накопичення цієї сполуки є недоліком для переробки цукрових буряків, оскільки погіршується процес вилучення цукру.
Кореляція вмісту інвертного цукру з даними експресії всіх досліджуваних сортів та часових точок за допомогою аналізу генної коекспресії (WGCNA) та подальшого функціонального аналізу показали, що більшість корельованих генів були пов'язані з захистом та реакцією на стрес. Це, головним чином, викликано позитивним регулюванням генів погано придатних до зберігання сортів під час Т4. Експресія генів, сильно корельованих із втратою цукру, збільшується під час зберігання дещо швидше у погано придатних до зберігання сортах. Як виявляється, ці гени беруть участь у регуляції росту та розвитку клітин. Гени у модулях, які негативно корелюють з інвертним цукром та втратою цукру, показали, що не всі гени, пов'язані зі стресом та захистом, мають вищий рівень експресії у поганих сортах через 13 тижнів зберігання; значна їх кількість має кращу експресію у хороших сортах, які можуть включати важливі гени абіотичного стресу (наприклад, дефіциту води) та стійкості до патогенів.
Те, що механізми захисту від патогенних мікроорганізмів активувались переважно в погано придатних до зберігання сортах, підтверджується аналізом, який вказує на розвиток вірусних та грибкових збудників, що впливають на збереженість буряків. Відомо, що, зокрема, три з виявлених родів грибів (Alternaria, Botrytis та Colletotrichum) викликають імунну реакцію клітинної стінки рослин.
Гени-концентратори, пов'язані з придатністю цукрових буряків до зберігання
За допомогою WGCNA вдалося визначити декілька генів-концентраторів, які корелюють з придатністю цукрових буряків до зберігання та можуть діяти як маркерні гени передбачення. Серед шести генів-концентраторів, які виявилися високо корельованими з інвертним цукром, були три халконсинтази, ключові ферменти біосинтезу фавоноїдів, які, як відомо, індукуються в абіотичних та біотичних стресах і відіграють важливу роль у стійкості рослин, підтверджуючи гіпотезу про підвищену реакцію на стрес у погано придатних до зберігання сортах. Крім того, була виявлена кафеол-КоА-О-метилтрансфераза, яка також бере участь у біосинтезі фавоноїдів, але особливо у лігніноутворенні, що, ймовірно, натякає на біосинтез фенілпропаноїдів, викликаний впливом стресу, у погано придатних до зберігання сортах, внаслідок чого у даному дослідженні неможливо підтвердити більше лігніноутворення. Серед генів-концентраторів, які сильно корелюють із втратами цукру, було виявлено три, які є шаперонами або діють із ними. Двоє беруть участь у системі HSP70, яка допомагає білкам досягти своєї власної конформації або відновити функцію після неправильного скручування внаслідок різних стресових умов. Іншим геном-концентратором втрати цукру є цитозольна триозофосфатізомераза, яка, як вважається, модулює виробництво АФК, як механізм стійкості.
|
Висновки На основі порівняльної транскриптоміки, анатомії цукрового буряка, даних про цукор та стандартні аналіти шести сортів буряків вдалося визначити ключові фактори, що впливають на придатність цукрових буряків до зберігання (визначається як втрати цукру та накопичення інвертного цукру під час зберігання). Окрім загального впливу, звузили генотипові відмінності до зберігання та під час періоду зберігання, тривалістю 13 тижнів. Сорти, які до зберігання мали більшу кількість клітин паренхіми та камбіальних кілець, а також тоншу перидерму, показали кращу поведінку під час зберігання. Крім того, негативне регулювання генів, що беруть участь у процесах дозрівання, здавалося потенційною передумовою для гарного зберігання, а також позитивного регулювання специфічної, очевидно, більш ефективної системи захисту від патогенів. Однак після 13 тижнів зберігання хороші сорти, здавалося, краще справлялися з гіперосмотичним станом клітин, демонструючи негативне регулювання клітинного дихання та вуглеводний метаболізм, а також менший захист та реакцію на стрес. Цікаво, що в добре придатних до зберігання сортах було виявлено вищий вміст альфа-аміно азоту, що вказує на осмозахисну функцію під час зберігання. Врешті-решт, гени-концентратори, які корелюють з придатністю цукрових буряків до зберігання, можуть додатково діяти як маркерні гени передбачення. |