Фізіологічна та метаболічна характеристики молодих цукрових буряків в умовах посухи (частина 3)

За матеріалами наукової статті на тему «Фізіологічна та метаболічна характеристики молодих цукрових буряків в умовах тимчасової посухи» («A physiological and metabolic characterization of young sugar beets under temporary drought»), Боннський університет, Німеччина, 2018 р.
Метаболомічний аналіз в умовах тимчасової посухи
У регіонах з періодичними посушливими періодами урожайність цукрових буряків залежить від здатності рослини до відновлення після припинення дії стресу. Було визначено відмінності в метаболічному відновленні пагонів та коренів цукрових буряків після посухи з високою тимчасовою роздільною здатністю. Рослини піддавалися впливу посухи протягом 13 днів, після чого починався період відновлення зрошення, який тривав 12 днів. Посуха спровокувала зміни в первинному метаболізмі, особливо збільшення амінокислот в обох органах рослин, але пагони та корені по різному реагували на відновлення зрошення. Після тимчасової нормалізації більшості метаболітів за 8 днів, повторне накопичення амінокислот у пагонах може свідчити про стресовий слід, сприятливий для майбутніх посушливих періодів. Механізми відновлення виявилися важливими під час початкового відновлення і здійснювалися за рахунок росту щонайменше 12 днів. Ці результати свідчать про те, що реакції відновлення метаболізму можуть бути пов'язані з певними функціями та супутніми різними рівнями стресу в наземних та підземних органах рослин. Що стосується метаболізму, його відновлення не було простим припиненням стресових реакцій.
Стабільна урожайність в мінливих умовах доступності води має стратегічне значення для забезпечення продовольством постійно зростаюче населення світу. Незважаючи на те, що за останні 100 років в Європі щорічна кількість опадів незначно змінилася, метеорологи спостерігають більший зсув між сезонами, тобто більш тривалий посушливий період навесні та влітку. Отже, сільськогосподарські культури відчувають все більше труднощів у боротьбі зі зміною умов навколишнього середовища за високу врожайність. Щодо виробництва цукрових буряків, на період 2021-2050 років передбачається скорочення виходу цукру через вплив посухи приблизно на 1 т цукру га-1 на півночі Франції, Бельгії та заході/центрі Польщі. Хоча часто описується вплив довготривалої посухи на фізіологічні та метаболічні процеси рослин, дослідження метаболічних реакцій рослини на відновлення зрошення обмежені. Швидке відновлення після посушливих періодів є бажаною рисою культури, тим більше, що рослини, як правило, піддаються неодноразовим посушливим періодам протягом свого життєвого циклу, які можуть набувати серйозного характеру.
Відновлення визначає період часу після припинення дії стресу до встановлення нової фізіологічної та метаболічної рівноваги і є вирішальним етапом у метаболізмі рослин. У відповідь на стрес фізіологічні адаптації та модифікації метаболізму призводять до накопичення метаболітів, у тому числі захисних сполук, які можуть надавати стійкість та резистентність до стресу від посухи. Після того, як стрес припиняється, починається процес відновлення, і рослина повинна досягти балансу між вкладенням ресурсів у зменшення пошкоджень, підтримуючи акліматизацію (підготовку до майбутніх стресових періодів), або у відновлення росту (переналаштування). Якщо переналаштування максимізує ріст та врожайність в сприятливих умовах, виникає ризик серйозних та, можливо, фатальних ушкоджень, якщо стрес повториться. З іншого боку, підтримка акліматизації робить рослину «насторожі» майбутніх стресових періодів (залишає так званий стресовий слід), але за рахунок зменшення росту чи розвитку та скорочення врожайності. За останніми даними, такий «стресовий слід» є досить рідкісним і повернення до початкового (до початку стресу) метаболічного та фізіологічного стану є більш поширеним явищем, але метаболічні дослідження, що підтверджують цю гіпотезу, все ще досить обмежені. Можливо, проміжні форми відновлення (певною мірою, але не до початкового рівня) можуть бути більш поширеними, оскільки вони представлятимуть найбільш перспективну стратегію реагування, принаймні, у регіонах, де періодичні стреси, як правило, нестабільні та непередбачувані.
Під час процесу відновлення обмінна енергія переходить у підготовку та налагодження відновлення фотосинтезу та дихання, високосинхронізовані та складні процеси. Щодо цукрових буряків, доступні дослідження процесів відновлення після посухи в основному обмежуються описом змін біохімічного складу та накопичення сахарози в корені або впливом тимчасової та тривалої посухи на урожайність, фотосинтез та накопичення вуглецю. Для підтримки високої врожайності особливе значення має швидке відновлення пагонів та коренів після посухи для забезпечення поглинання води, поживних речовин та продовження період накопичення сахарози. Потрібне краще розуміння подібності та особливостей пагонів та коренів у метаболічному регулюванні та відновленні після тимчасової посухи, що є головною метою даного дослідження.
Дана робота спрямована на виявлення та характеристику основних метаболітів первинного метаболізму з метою розкриття метаболомічної стратегії цукрових буряків в умовах тимчасового стресу. Комплексне використання метаболомічних інструментів, таких як спектроскопія протонним ядерним магнітним резонансом (1H NMR) та системна біологія, є потужним інструментом для всебічного огляду задіяних шляхів та виявлення важливих сполук метаболічної реакції. Оскільки рослинні метаболіти надзвичайно різноманітні як за своєю біологічною функцією, так і за хімічною будовою, аналіз 1H-NMR є прекрасним інструментом для вивчення не лише складу сполук рослинного метаболізму, а й усіх динамічних аспектів.
Результати
Подолання рослинами порушень водного режиму та мембранної стійкості, викликаних впливом посухи
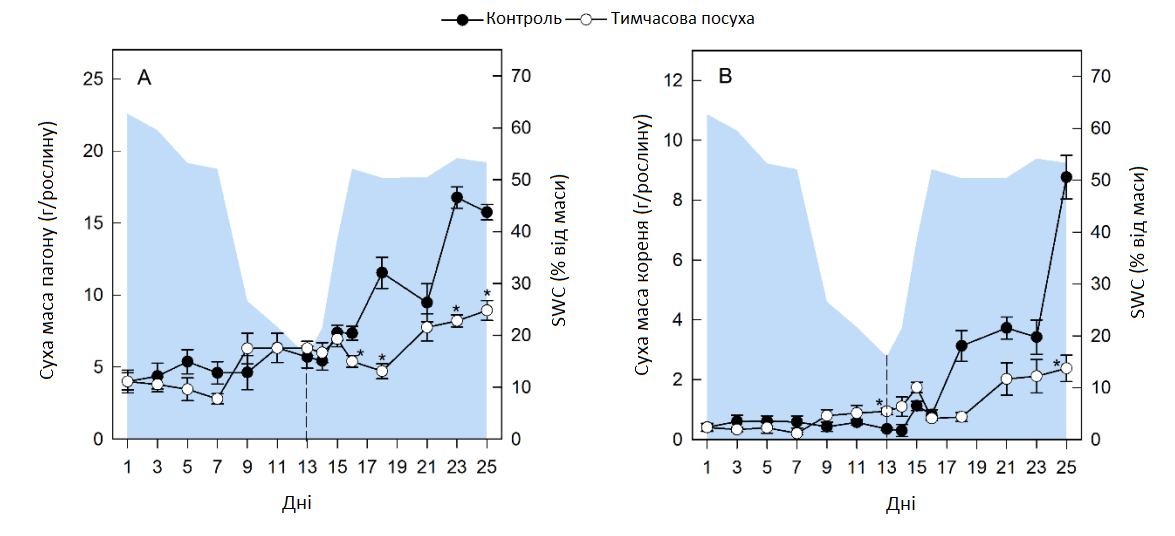
Під час тривалої посухи вміст води в ґрунті (SWC) повільно знижувався протягом перших 7 днів, а потім швидше до 13 дня (Графік 2). За цих умов суха маса пагону (DW) не значно зменшилася, порівняно з контрольними рослинами, які повільно розвивалися з BBCH 16-17 (6-7 листочків, 1 день) до BBCH 17-19 (7-9 листочків, 25 день), але рослини, що зазнали впливу посухи, мали значно вищу суху масу коренів на кінець періоду посухи (Графік 2).
Графік 2. Зміна біомаси пагонів та коренів цукрових буряків. Суха маса пагонів (А) та коренів (В) контрольних рослин (темні кола) та рослин під впливом посухи (білі кола). Зрошення було відновлено через 13 днів, як показано пунктирною лінією. Графік являє собою ваговий вміст ґрунтової води на основі субстрату (SWC, % від маси). Усі значення — середні арифметичні ± (n = 4). Значні відмінності контрольних рослин (α = 0,5) позначаються як * P < 0,05.
Через 7 днів відносний вміст води (RWC, Графік 3, A) значно зменшився та досягнув мінімального значення 37 ± 2% на 11 день посухи, тоді як найбільше зниження осмотичного потенціалу (ОР, Графік 3, B) спостерігалося між 9 та 13 днями посухи з кінцевими значеннями 1,56 ± 0,2 МПа. Було виміряно витік електроліту (EL, Графік 3, С) та малондіальдегід (MDA, Графік 3, D), які є показниками пошкодження мембрани внаслідок перекисного окислення ліпідів, а перші ознаки пошкодження мембрани в пагоні спостерігалися через 7-9 днів від початку посухи.
Графік 3. Параметри водного режиму рослини та пошкодження мембрани. Відносний вміст води (RWC, A), осмотичний потенціал (OP, B), витік електроліту (EL, C) та концентрації малондіальдегіду (MDA, D) пагону в умовах регулярного зрошення (контролю, темні кола) та тимчасової посухи (білі кола). Графік являє собою ваговий вміст ґрунтової води на основі субстрату (SWC, % від маси). Зрошення було відновлено через 13 днів, як показано пунктирною лінією. Усі значення — середні арифметичні ± (n = 4). Значні відмінності контрольних рослин (α = 0,5) позначаються як * P < 0,05.
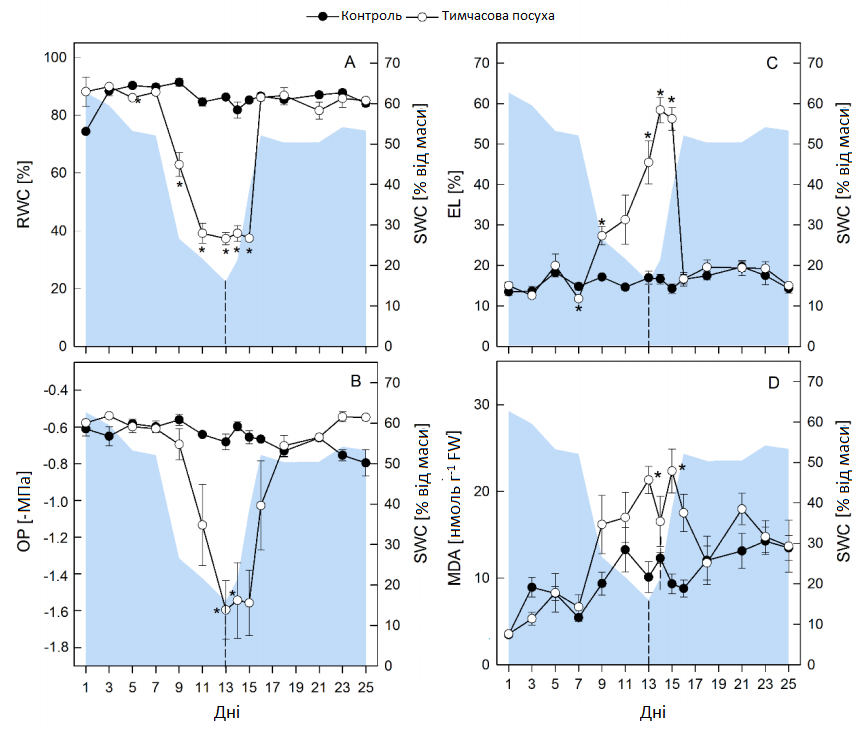
Після початку відновлення зрошення молодші листочки відновили тургор протягом 2 днів, але найстаріше листя не змогло повністю відновитися до кінця всього експерименту (Графік 4). Період затримки 5 днів спостерігався до початку відновлення росту рослин, які постраждали від посухи, але зберігався низький відносний темп росту лише 26% (пагони) та 31% (корені), порівняно з контрольними рослинами, протягом 14-25 днів. І відносний вміст води (RWC), і осмотичний потенціал (OP) показали затримку в 2 дні після відновлення зрошення, перш ніж почали відновлюватися, а потім досягли контрольних рівнів за 1 (RWC) та 2 дні (OP) відповідно (Графік 3, A, В). Концентрації малондіальдегіду (MDA) повернулися до контрольного рівня за 4-6 днів. Однак витік електроліту (EL) продовжував збільшуватися 1-2 дні в період відновлення зрошення, але потім відновився швидше, ніж MDA, та досягнув контрольного рівня за 3 дні (Графік 3 C, D).
Графік 4. RGB-зображення цукрових буряків, що постраждали від тимчасової посухи. Молоді рослини Beta vulgaris, що перебувають в умовах регулярного зрошення (контролю) на 13 (A), 15 (C) та 25 (E) день експерименту, рослини під дією стресу від посухи (B) та в умовах відновлення зрошення (D, F).

Тимчасова посуха призводить до змін у первинному метаболізмі
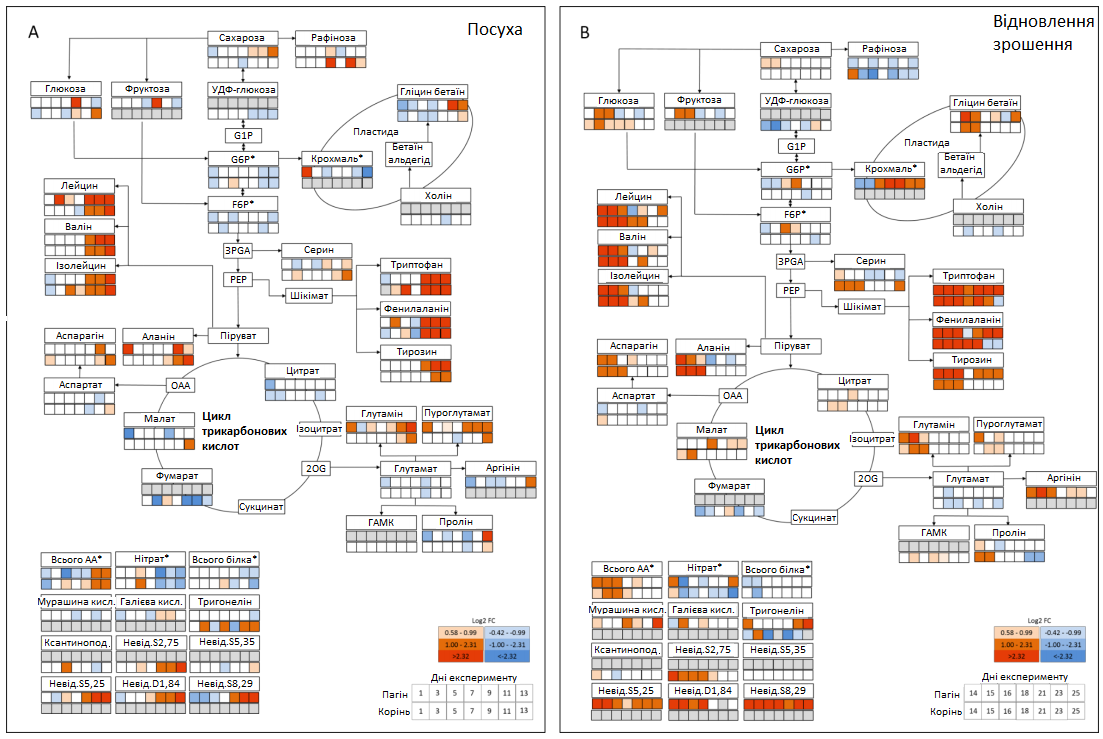
Загалом за допомогою 1H-NMR було виявлено 30 метаболітів, у тому числі п’ять вуглеводів, 17 амінокислот (АА), п'ять органічних кислот, два четвертинного з’єднання амонію та один алкалоїд (Таблиця 1, Графік 5). Для порівняння змін, викликаних посухою, (1-13 дні) та відмінностей між пагонами та коренями (13-25 дні) під час відновлення зрошення, було побудовано схеми метаболізму, що показують зміну між добре зрошуваними рослинами та рослинами під дією стресу від посухи (Графік 6).
Таблиця 1. Хімічні зрушення, що використовуються для ідентифікації та кількісного визначення метаболітів у спектрі 1H-NMR. Хімічні зрушення коренів цукрових буряків та полярні екстракти листя (у D2O), виражені у відносних значеннях до резонансу TSP при 0 мг/м3. s: синглет, d: дублет, dd: подвійний дублет, t: триплет, m: мультиплет.
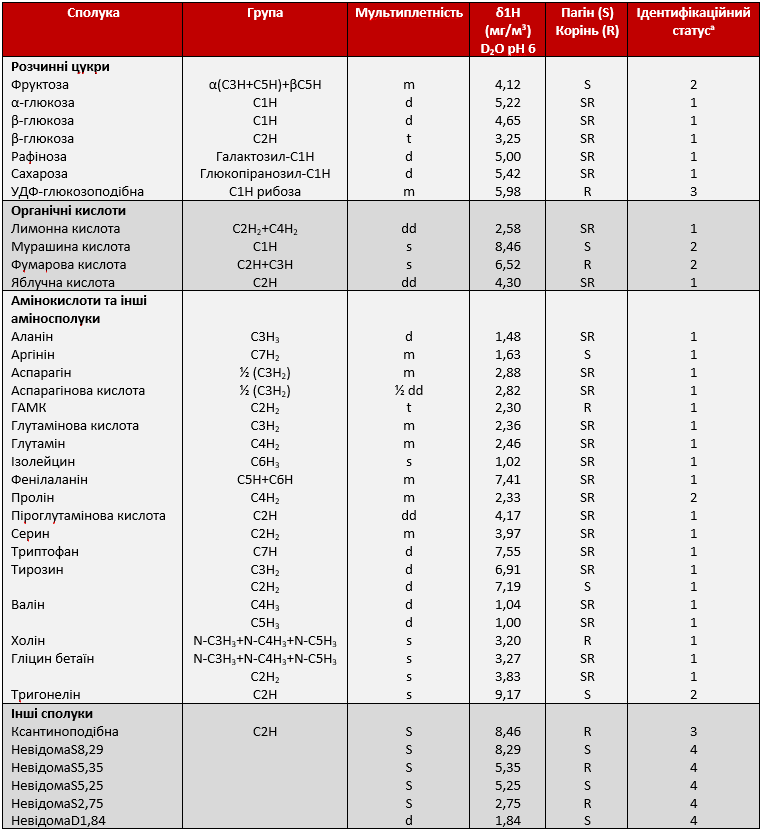
a Рівень ідентифікації відповідно до MSI: 1, Ідентифіковані сполуки (перевірені стандартом); 2, Імовірно анотовані сполуки; 3, Імовірно характеризовані класи сполук; 4, Невідомі соплуки.
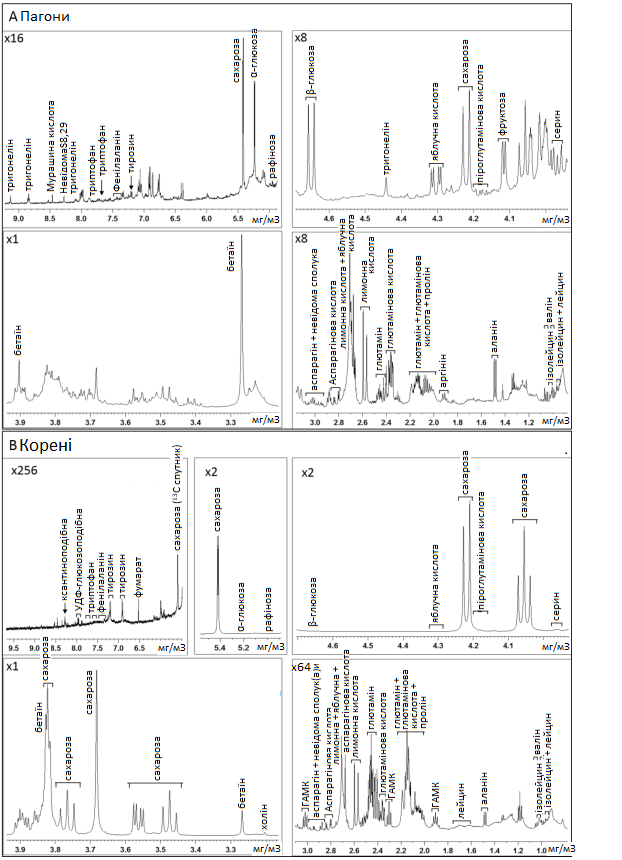
Графік 5. Репрезентативні спектри 1H-NMR пагонів та коренів цукрових буряків. Два репрезентативних спектри 1H-NMR у різних збільшеннях полярних екстрактів пагонів (A) та коренів (B) добре зрошуваних рослин Beta vulgaris на 15 день експериментального періоду. Цифри у лівому верхньому кутку відповідають збільшенню.
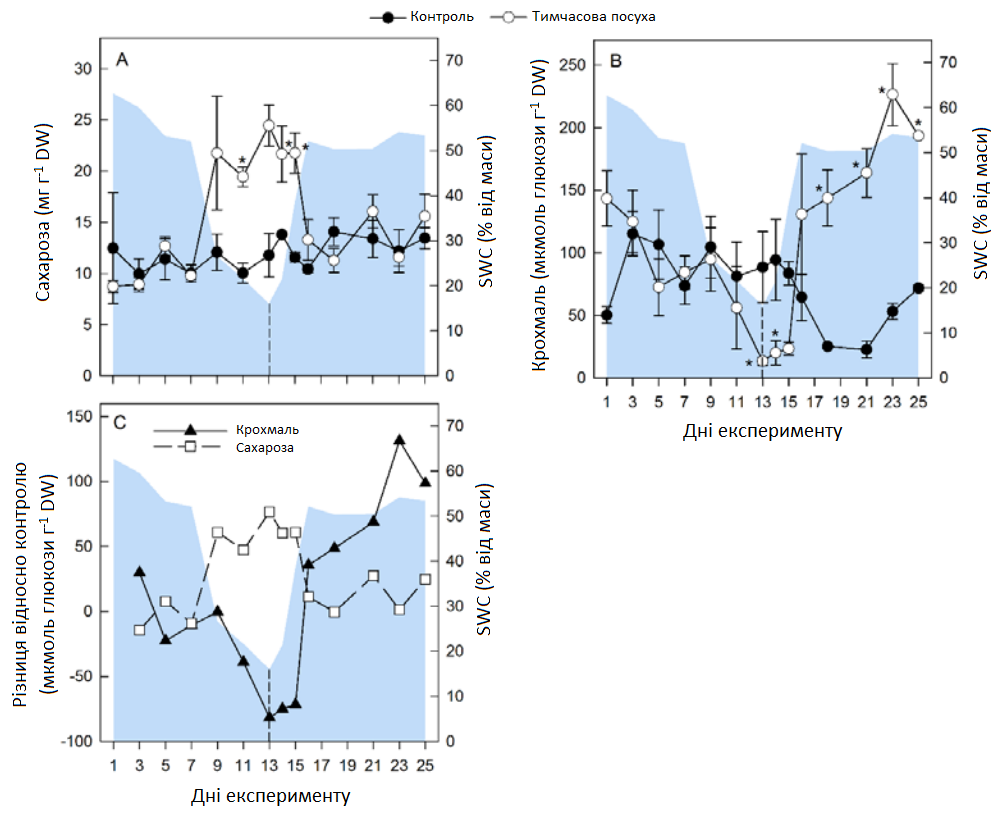
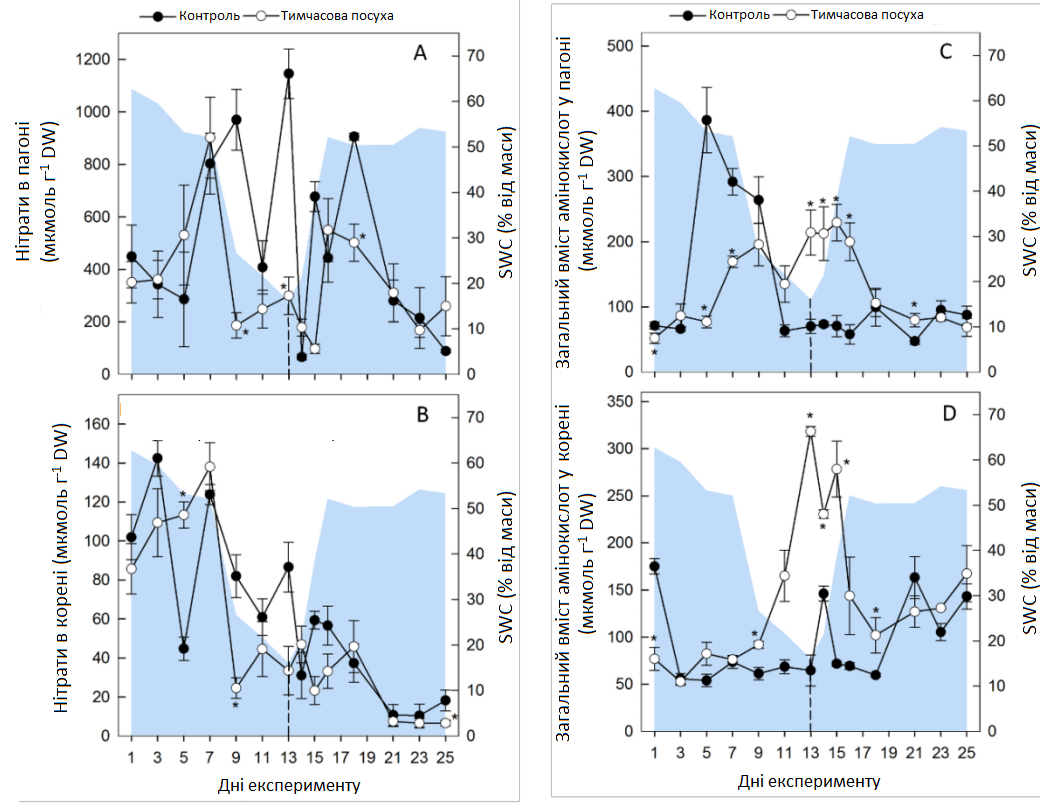
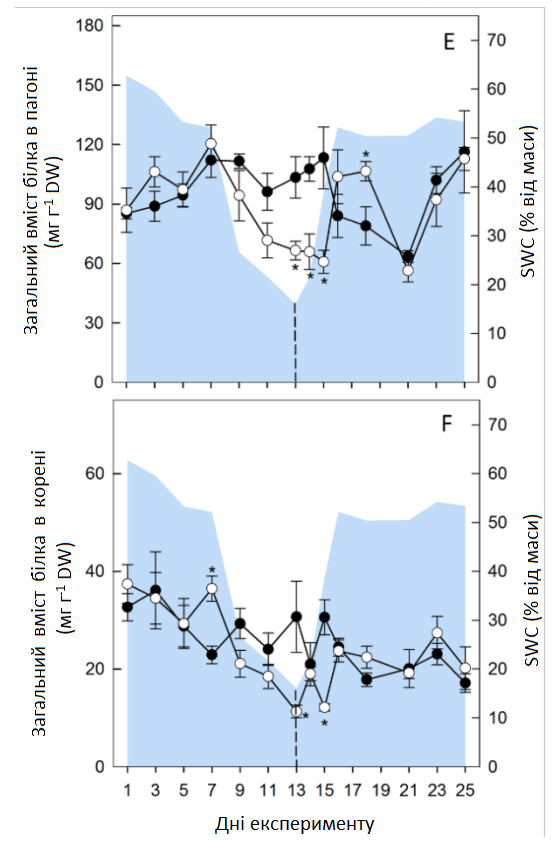
Тимчасова посуха викликала зміну декількох метаболітів, включаючи цукри, органічні кислоти, сумісні осмоліти та особливо амінокислоти (АА) в обох органах рослини (Графік 6 А). У листі спостерігалися протилежні ефекти для сахарози та крохмалю, де рівень сахарози збільшувався, а концентрація крохмалю знижувалася до кінця посушливого періоду (Графік 6 А; Графік 7). Кількісно вимірювані проміжні продукти циклу трикарбонових кислот (цитрат, малат, фумарат) практично не зазнали змін під час посухи, за винятком зменшення рівня фумарату в коренях (Графік 6 А). Перепрограмування метаболізму, спричинене посухою, призвело до збільшення загальної кількості амінокислот (AAt), а також до зменшення нітратів та білка (Графік 8). Найінтенсивніше збільшення спостерігалося для амінокислот із розгалуженими ланцюгами (BCAA: лейцину, ізолейцину, валіну), аланіну, похідного пірувату, та ароматичних амінокислот (ААА: триптофану, фенілаланіну, тирозину), похідних фосфоенолпірувату, при цьому максимальна зміна спостерігалася для фенілаланіну (> 200 разів у пагонах, > 70 разів у коренях). Особливо в пагонах, спричинене посухою збільшення глютаміну, піроглутамату, аргініну та проліну було пов’язано зі зменшенням їхнього попередника — глутамату (Графік 6 А). Відповідно, збільшення аспарагіну в пагонах супроводжувалося зменшенням його попередника — аспартату. Четвертинне з'єднання амонію гліцин бетаїн (GB) та пролін також накопичувалися в пагонах до кінця стресового періоду та лише незначно в коренях, де збільшення GB все-таки було пов’язане зі зменшенням його попередника — холіну (Графік 6 А). Загалом, карта метаболічного шляху вказує на те, що в умовах посухи гліколіз та цикл трикарбонових кислот були досить регульованими, тоді як рівні амінокислот (АА) були значно підвищені.
Графік 6: Метаболічна карта стресів від посухи та відновлення зрошення цукрових буряків. Відмінності метаболомічного профілю 1H-NMR протягом 13 днів посухи та відновлення зрошення. (А) Посуха, 1-13 дні, (В) відновлення зрошення, 14-25 дні. Верхній рядок вказує на метаболічні зміни в пагоні, нижній — на зміни в корені. Червоний та синій кольори вказують на надлишок (log2 Fold change, log2FC) кожного метаболіту відносно контрольних рівнів. Сірий колір вказує на те, що відповідний метаболіт не був ідентифікований (лише в пагоні: фруктоза, крохмаль, мурашина кислота, галієва кислота, невідомаS5,25, невідомаD1,84. Лише в корені: УДФ-глюкозоподібні цукри, холін, ГАМК, ксантиноподібний метаболіт, невідомаS2,75, невідомаS5,35). Зірочки (*) вказують на те, що метаболіти визначалися за допомогою роботизованих ферментних аналізів (G6P, F6P, крохмаль, загальні амінокислоти (всього АА, нітрати, білок).
Графік 7. Зміни концентрацій сахарози та крохмалю. Зміни сахарози (А) та крохмалю (В) в контрольних умовах (темні кола) та в умовах тимчасової посухи (світлі кола). (C) різниця в концентраціях крохмалю (темні трикутники) та сахарози (світлі квадрати) в умовах тимчасової посухи по відношенню до контрольних умов. Усі значення — середні арифметичні ± (n = 4). Значні відмінності від контрольних рослин (α = 0,5) позначаються як * P < 0,05.
Графік 8. Цільовий аналіз нітратів, загального вмісту амінокислот та білка. Зміни нітратів (A, пагін; B, корінь), загального вмісту амінокислот (C, пагін; D, корінь) та загального білка (E, пагін, F, корінь) в контрольних умовах (темні кола) та в умовах тимчасової посухи (світлі кола). Усі значення — середні арифметичні ± (n = 4). Значні відмінності від контрольних рослин (α = 0,5) позначаються як * P < 0,05.
 Різна динаміка пагонів і коренів під час процесу відновлення
Різна динаміка пагонів і коренів під час процесу відновлення
Більшість підвищених метаболітів під час посухи наблизилися або повернулися до контрольного рівня протягом 15-18 днів (2-5 днів після відновлення зрошення, DAR), за винятком крохмалю, який збільшувався протягом усього періоду відновлення і досягнув значно більших значень на кінець експерименту, порівняно з контрольним рівнем (Графік 6). Деякі чіткі відмінності спостерігалися між пагонами та коренями (Графік 6 B). На початку періоду відновлення зрошення в пагонах глюкоза (та аналогічно фруктоза) швидко зменшилися нижче контрольних рівнів, але потім показали другий максимальний рівень протягом 15-16 днів, тоді як у коренях глюкоза поверталася до контрольних рівнів досить повільно (23 день).
У коренях та пагонах концентрації цитрату та малату незначно збільшилися під час відновлення зрошення, тоді як фумарат (виявлений лише в коренях) залишався нижче контрольного рівня протягом усього періоду відновлення. Відновлення зрошення зупинило викликане посухою збільшення амінокислот у пагонах протягом 5 днів (18 день), тоді як у коренях накопичені амінокислоти зменшувалися повільніше, але постійно, та досягли контрольних рівнів на 23 день. Помітне накопичення амінокислот та менш виражене лейцину, валіну та гліцин бетаїну показали друге сильне збільшення напередодні останніх днів періоду відновлення зрошення в пагонах, але не в коренях.
Основною відмінністю коренів та пагонів була реакція серину, яка була сильнішою в коренях, де він повільно відновлювався до контрольних рівнів під час відновлення зрошення, а також демонстрував сильне друге збільшення напередодні завершення експерименту, подібно до динаміки, що спостерігалася з накопиченням амінокислот у пагонах. Підсумовуючи вище сказане, можна сказати, що динаміка зміни метаболітів вказує на чіткі зміни метаболічної активності між пагонами та коренями в умовах посухи та відновлення.
Із метою пошуку метаболітів, які були найважливішими індикаторами стресу та відновлення, а також для оцінки того, чи та як змінилися викликані посухою умови під час відновлення зрошення, проводився аналіз основних компонентів (PCA) у пагонах та коренях із використанням матриці, що містить дані про 27 (пагони) та 26 (корені) кількісно оцінених метаболітів у 84 зразках кожного пагону та кореня (Графік 9). PCA дозволив виявити метаболіти, що беруть участь у динамічній реакції.
Щодо PCA пагону (Графік 9 A, B), перші два основні компоненти пояснювали 53,6% загальної мінливості, при цьому 40% припадало на перший компонент (PC1) та 13,6% — на другий компонент (PC2). У графіку розрахунків (Графік 9 А) PC1 відокремлював велику групу, що містить контрольні зразки рослин, які пережили слабкий стрес (< 9 днів) та пізній початок зрошення (> 15 днів), від зразків, взятих на пізніх стадіях посухи та на початку відновлення зрошення (9, 15 день), що свідчить про те, що PC1, здається, пов'язаний з інтенсивністю стресу. PC2, як правило, відокремлював молодші рослини (~1-15 день, негативна сторона) від старих рослин (~16-25 день, позитивна сторона), що дозволяє припустити, що PC2 пов'язаний з розвитком пагону. Однак це розділення було менш чітким, ніж для PC1, відповідно до більш поступової зміни метаболізму протягом усього періоду розвитку. Траєкторії (Графік 9 А) візуалізують відмінності в зміні метаболізму контрольних рослин, рослин в умовах посухи та відновлення зрошення.
Порівняння графіків розрахунків (Графік 9 А) та навантажень (Графік 9 B) показало, що зразки, взяті на пізніх стадіях посухи та раннього відновлення зрошення, мали тенденцію до більш високого вмісту амінокислот (АА), включаючи тирозин, триптофан, фенілаланін, піроглутамат, лейцин, ізолейцин, валін та глютамін. Поряд із PC2, молодші рослини, здавалося, характеризувалися вищим вмістом тригонеліну, а старіші — більш високим вмістом цитрату.
Щодо PCA кореня (Графік 9 C, D), перші два основні компоненти пояснювали 68,3% загальної мінливості (PC1 — 50,6%, PC2 — 17,7%). У графіку розрахунків (Графік 9 С) PC1 відокремлював одну велику групу, що містить рослини, які пережили слабкий стрес (< 11 днів) та пізній початок зрошення (> 16 днів), від меншої групи, що характеризується зразками, відібраними на пізніх стадіях посухи та раннього відновлення зрошення (11-16 день), що свідчить про те, що PC1 здається пов'язаний з інтенсивністю стресу. PC2 прагнув поступово відокремлювати зразки на ранніх стадіях від зразків на пізніх стадіях розвитку. Порівняння графіків розрахунків (Графік 9 С) та навантажень (Графік 9 D) показало, що зразки, взяті на пізніх стадіях посухи та раннього відновлення зрошення, мали більш високий вміст амінокислот, подібно до листя, а також сахарозу та гліцин бетаїн. На пізніх стадіях коренеплоди мали більший вміст аспартату, глутамату та рафінози.
Графік 9. Розрахунки PCA та навантаження цукрових буряків, що пережили посуху. Результат аналізу основних компонентів (PCA) профілів 1H-NMR. Графік розрахунків пагону (A) та кореня (C), графік навантаження пагону (B) та кореня (D); розмір зразка n = 3. Графіки PC1 x PC2 становлять 53,6% і 68,3% від загальної дисперсії для пагонів і коренів відповідно. На графіку розрахунків кола представляють контрольні рослини, а трикутники —рослини в умовах тимчасового стресу. Траєкторії на графіку розрахунків представляють часовий розвиток метаболічної реакції під час експерименту (синя стрілка — контроль; червона — посуха, зелена — відновлення зрошення). Asn, Asp — аспарагін; Gaba — γ-амінобутанова кислота; GB — гліцин бетаїн; Gln — глутамін; Glu — глутамат; Ile — ізолейцин; Leu — лейцин; Val — валін; Phe — фенилаланін; Pro — пролін; Pyro-Glu — піроглутамат; Ser — серин; Trp — триптофан; Tyr — тирозин.
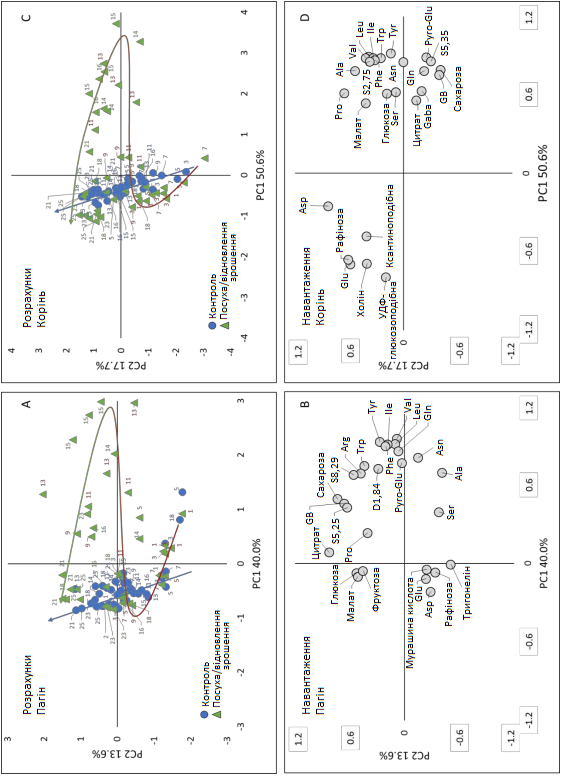
Аналіз основних компонентів (PCA) як коренів, так і пагонів підтверджує, що основні метаболічні зміни, викликані стресом, відбулися протягом останніх 3-5 днів посухи і тривали ще 3 дні в період відновлення, що відображає реакцію водного режиму (RWC, OP) і пошкодження мембрани (EL, MDA) під час стресу та фази відновлення. Після відновлення зрошення зразки обох органів показали зворотну траєкторію (Графік 9 А, С) та повернулися до контрольних рівнів протягом 16-17 днів, що вказує на тимчасовий характер метаболічних змін, викликаних посухою. Однак зразки коренів, взяті протягом 23-25 днів, мали тенденцію знову відокремлюватися від контрольних рослин у плані першого основного компоненту (PC1, Графік 9 С), що дозволяє припустити, що через 10 днів після відновлення зрошення (DAR, 23 день) корені метаболічно відрізнялися від контрольних умови, тоді як у пагонах цього не було.