Фізіологічна та метаболічна характеристики молодих цукрових буряків в умовах посухи (частина 1)

Цукровий буряк (Beta vulgaris) — головна цукроносна культура помірного кліматичного поясу Європи, яка має високе економічне значення. На цукрові буряки припадає 25% світового виробництва цукру. Крім того цукор-сирець відіграє важливу роль у хімічній промисловості, а побічні продукти переробки цукрових буряків, такі як буряковий жом, використовуються як корм для ВРХ, а також служать основою для багатьох інших продуктів. Крім того цукровий буряк почали використовувати для виробництва біогазу завдяки його хорошим ферментаційним властивостям, порівняно з іншими енергетичними культурами.
Дворічна культура, яка належить до родини Амарантових (раніше Лободові) походить з прибережних районів Північного моря. Найдавніші знахідки солестійкої рослини відносяться до епохи Неоліту, які були виявлені на півночі Голландії. Але використання знайдених залишків буряка не зовсім зрозуміле, ймовірно, для споживання використовувалося лише листя. У 16 столітті було виявлено, що екстракт м’якоті цукрових буряків виділяє солодкий сироп, а потім у 1747 році хімік Андреас Сигізмунд Маргграф показав, що кристали цукру, отримані з кормового буряка (Runkelrübe, B. vulgaris) з вмістом цукру лише 4% хімічно ідентичні кристалам, одержаним із цукрової тростини (Saccharum officinarum). Оскільки в Runkelrübe містилося менше цукру, плантатори намагалися збільшити вміст цукру шляхом селекції. У 1802 році в Кунерні (Пруссія) Франц Карл Ашард збудував перший цукровий завод. Незважаючи на те, що головним проривом для цукрових буряків стала Континентальна блокада Наполеона (1809-1814), низький вміст цукру 8% все ще був причиною імпорту цукрової тростини. Систематична селекція в наступні століття призводить до постійного збільшення вмісту цукру в буряках. У сучасних сортах буряків вміст цукру досягає 17-22% (від свіжої маси), а цукровий буряк все ще залишається рослиною з найвищим вмістом цукру в цукровій галузі Європи.
За матеріалами наукової статті на тему «Фізіологічна та метаболічна характеристики молодих цукрових буряків в умовах тимчасової посухи» («A physiological and metabolic characterization of young sugar beets under temporary drought»), Боннський університет, Німеччина, 2018 р.
Незважаючи на широкі та постійні зміни в європейській цукровій галузі протягом останніх років та на світовому ринку цукру, вирощування цукрових буряків все ще є основою для виробництва цукру в Німеччині вже більше 200 років. У 2015/2016 МР в Європі було вироблено приблизно 14,9 млн т цукру, де Франція є найбільшим виробником із урожайністю 12,9 т/га, за нею слідують Німеччина та Польща з 11,3 т/га та 8,5 т/га відповідно (WZV та VdZ, 2016).
У зв’язку з екологічними та кліматичними змінами останніх десятиліть, необхідні нові підходи до селекції та зміни потреб у вирощуванні. Таким чином, стратегії селекції спрямовані не лише на збільшення виходу цукру. Через зміни атмосферних опадів та температурного режиму, селекція та дослідження фокусуються на проблемі посухи та холодостійких сортів. Крім того стратегії селекції також залишаються орієнтованими на стійкість до ризоманії, церкоспорозу та нематоди. Що стосується вдосконалення технологій механізаваного збирання врожаю, автоматизованого внесення добрив та захисту рослин за допомогою оптичних датчиків або камер, селекція у контексті розвитку різних форм пагону та головного кореня цукрового буряка продовжує відігравати важливу роль. Усі ці фактори демонструють різноманітність старих та нових селекційних ознак. Таким чином, селекція спрямована на одночасну оптимізацію декількох складних ознак, які, крім того, знаходяться під полігенним контролем. Вміст цукру, технічна якість сировини та покращення виділення біомаси є лише декількома параметрами, але для подальшої селекції потрібен пошук різноманітних фенотипічних та генотипних ознак.
Дефіцит води у виробництві цукрових буряків
Достатня кількість опадів протягом періоду вегетації, з червня по вересень, є основою для ефективного росту та розвитку цукрових буряків. Завдяки своїм солестійким особливостям, цукрові буряки є досить стійкими до посухи, особливо, в порівнянні з іншими культурами весняного посіву, адже завдяки відносно хорошій ефективності поглинання води вони можуть досить добре витримувати посуху протягом певного періоду часу. За допомогою глибокої та широко розгалуженої кореневої системи, яка може досягати глибини до 110 см, цукровий буряк може «видобувати» та засвоювати велику кількість ґрунтової води. Однак його здатність регулювати транспірацію обмежена, тому в’янення є одним із перших симптомів, який можна спостерігати майже регулярно під час тимчасових посушливих періодів, що часто поєднуються з високим рівнем сонячного випромінювання, наприклад близько полудня влітку. Але це явище спостерігається також при регулярному водопостачанні, отже в'янення листя цукрових буряків є не тільки наслідком посухи, але й також ознакою недостатнього засвоєння води буряком, навіть при регулярній подачі води. Попри те, що цукровий буряк відновлюється після посухи, здатність рослин справлятися зі зменшенням дефіциту ґрунтової води обмежена, а водний потенціал листя знижується швидше, ніж потенціал ґрунтових вод. У помірному кліматичному поясі річна кількість опадів приблизно 600 мм мають вирішальне значення для високого виходу цукру з найбільшою кількістю опадів протягом другої частини сезону (червень-вересень). Однак це не є загальною тенденцією, особливо під час довготривалих літніх періодів на півдні Європи. Тут вирощування цукрових буряків можливе лише в умовах зрошення. Але в таких основних виробничих країнах, як Росія, Україна, Польща, Німеччина та Англія, де зазвичай спостерігається достатня кількість опадів протягом року, втрати врожаю через посуху можуть досягати 15-40%.
Краще розуміння фізіологічних та метаболічних змін під час періодичних або постійних посушливих періодів покращить усталені підходи у виведенні посухостійких культурних сортів, зробить внесок у розробку нових стратегій селекції та запропонує нові варіанти критерій відбору для стійкості до посухи. Оскільки механізми адаптації в умовах посухи подібні до інших абіотичних стресів, таких як стрес від холоду або сольовий стрес, стійкість до посухи також збільшує стійкість до інших стресів. Для досягнення цих цілей потрібна попередня і винятково диференційована картина змін, викликаних посухою, що може бути досягнуто поєднанням класичних деструктивних методів, а також недеструктивних підходів, наприклад тепловізуалізації.
Процес відновлення
Процес відновлення описує період після припинення стресу до встановлення нового фізіологічного та метаболічного гомеостазу та є важливим етапом у метаболізмі рослин. У відповідь на стрес фізіологічні пристосування метаболізму дозволяють синтезувати метаболіти, включаючи захисні сполуки, наприклад цукри або четвертинні з'єднання амонію, які можуть забезпечувати стійкість до стресів, викликаних посухою. Коли стрес припиняється, починаються процеси відновлення, і рослина повинна досягти балансу між використанням ресурсів для зменшення пошкоджень підтримуваною аклімацією (підготовкою до майбутніх стресових умов) або новим відростанням/відтворенням. Коли переналаштування максимізує ріст та врожайність в сприятливих умовах, виникає ризик серйозних та, можливо, навіть фатальних ушкоджень, якщо стрес повториться. З іншого боку, аклімація робить рослину «підготовленою» до майбутніх стресових умов, але за рахунок зменшення росту чи розвитку та скорочення врожайності.
Під час процесу відновлення обмінна енергія переходить у підготовку та налагодження фотосинтезу, дихання та біосинтезу ліпідів, високосинхронізованих та чутливих процесів. Щодо В. vulgaris, наявні дослідження процесів відновлення після посухи в основному обмежуються описом зміни хімічного складу коренеплоду та накопичення цукру або боротьби з впливом тимчасової та повторюваної посухи на урожайність, фотосинтез та апробацію вуглецю. Однак, бракує даних про краще розуміння подібних та відмінних особливостей коренеплодів та пагонів у метаболічному налаштуванні та відновленні після посухи. У цьому є гостра потреба, оскільки швидкість відновлення коренеплоду після посухи має особливе значення для забезпечення високої врожайності, щоб гарантувати хороше поглинання води та живильних речовин із метою запобігання втрати врожаю, при цьому не погіршуючи накопичення цукру.
Фенотипування
Поточне вживання слова «феном» (із давньогрецької «phaino» — я з’являюсь та «týpos» — форма) стосується всього фенотипу. Фенотип описує склад спостережуваних характеристик певного індивіду. Щодо рослин, фенотип враховує ріст та розвиток, морфологію, стійкість, урожайність, а також фізіологічні та біохімічні характеристики. Фенотип організму є результатом експресії геному, на який можуть впливати зовнішні та внутрішні фактори, наприклад, вплив навколишнього середовища, абіотичний та біотичний (зовнішній) стрес, взаємодія між зовнішніми факторами і самим геномом, наприклад мутації, викликані абіотичним (внутрішнім) впливом. Отже, взаємодія між геномом та навколишнім середовищем визначає фенотипічну пластичність рослини. Фенотипування налічує тисячі років, яке спочатку інтуїтивно виконувалося виробником, який проводив обстеження свого поля протягом сезону. Сьогодні фенотипування проводять фермери, селекціонери, галузь сільського господарства та наукові установи, яке спрямовані не лише на досягнення високої врожайності, але й удосконалення способів вирощування для поліпшення бажаних ознак, наприклад ефективності використання води або біомаси, раннє виявлення абіотичного та біотичного стресу. Фенотипування — це дослідження, яке швидко розвивалося протягом останніх десятиліть. Фенотипування, яке почалося як спостереження виключно неозброєним оком, сьогодні підтримується різними складними та часто автоматизованими системами високої пропускної здатності у фенотипувальних установах науково-дослідних інститутів, безпілотних літальних апаратах (БПЛА), що використовуються для спостереження та моніторингу посівних площ, а також у галузі сільського господарства. Незважаючи на те, що в останні роки відбулася масштабна розробка неінвазивних методів спостереження зовнішнього фенотипу, аналіз та інтерпретація внутрішнього фенотипу, що включають фізіологічні, біохімічні та, таким чином, метаболічні зміни, які остаточно визначають зовнішній фенотип, «пасуть задніх». Для усунення цього розриву, безумовно, необхідно поєднувати зовнішній підхід із високодинамічними внутрішніми процесами на біохімічному та фізіологічному рівнях. Лише такий підхід може дати картину реального та цілісного фенотипу, який є результатом взаємодії генотипу з навколишнім середовищем.
Головною метою даного дослідження є фізіологічна та метаболічна характеристика молодих цукрових буряків в умовах тимчасової посухи та їх відновлення з особливим акцентом на відмінності реакцій пагонів та коренів.
Для цього були досягнуті наступні цілі:
І. Розробка надійної та відтворюваної системи проведення досліджень, яка дозволяє контролювати відтворення безперервної посухи та відновлення зрошення цукрових буряків в тепличних умовах, а також достатній та економічно ефективний аналіз фізіологічних та метаболічних змін.
ІІ. Дослідження хронологічного порядку фізіологічних та метаболічних змін молодих цукрових буряків в умовах безперервної посухи та відновлення зрошення, з особливим акцентом на водний режим рослин та осмотичну корекцію, оцінену інвазивними та неінвазивними методами.
ІІІ. Детальний метаболічний підхід із використанням незарядженого ядерно-магнітного резонансу (1H-NMR) та цільових метаболітів аналізує основні метаболічні механізми цукрових буряків в умовах тимчасової посухи та відновлення зрошення, акцентуючи увагу на відмінностях процесів відновлення пагонів та коренів.
Рослини
У всіх експериментах використовувалися цукрові буряки сорту Паулета, стійкого до нематоди та ризоманії, рекомендованого для проблемних посівних площ, що страждають від посухи та бур’янів (особливо переліски однорічної — Mercurialis annua L.), характеризується хорошим розвитком молодих рослин. Постачальником насіння став доктор Брітта Шульц компанії KWS Saat AG, м. Айнбек, Німеччина.
Фенологічний розвиток рослин
Фенологічний розвиток рослин визначали за шкалою BBCH, яка розрізняє 9 основних стадій росту та розвитку рослин (Таблиця 1).
Таблиця 1: Шкала BBCH (частини) фенологічного розвитку цукрових буряків


Етапи росту та розвитку рослин можна розділити на дві секції. Перші 4 стадії росту відносяться до першого року розвитку, від проростання (BBCH 0) до розвитку органів вегетативного розмноження, стадії розетки (BBCH 4), а наступні стадії росту позначають другий рік розвитку, від появи суцвіття (BBCH 5) до старіння рослини (BBCH 9).
Вирощування рослин
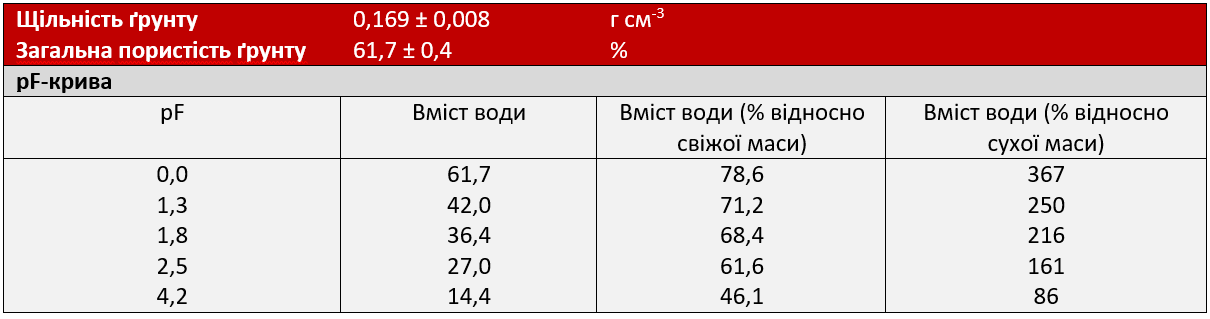
Рослини попередньо вирощувалися в горщиках, що містили суміш піщаного ґрунту (кварцовий пісок/ґрунт для розсади — 1:1). Рослини вирощувалися в тепличних умовах до появи та розвитку перших листочків. Під час вегетативного росту рослини вирощувалися в 2-літрових пластикових горщиках (11,3 х 11,3 х 21,5 см), наповнених 850 г субстратної суміші (70% білого торфу, 20% глинистого ґрунту, 10% перліту, Gepac, тип VM, Сінтал-Джосса, Німеччина). Властивості субстрату наведені на Графіку 1 та в Таблиці 2. Денна та нічна температури складали 24°C та 18°C при відносній вологості (RH) приблизно 75% і тривалості світлового дня 16 год з інтенсивністю світла > 250 мкмоль m-2 s-1 (Philips SON-T Agro 400W).
Графік 1: Властивості субстрату Gepac, тип VM
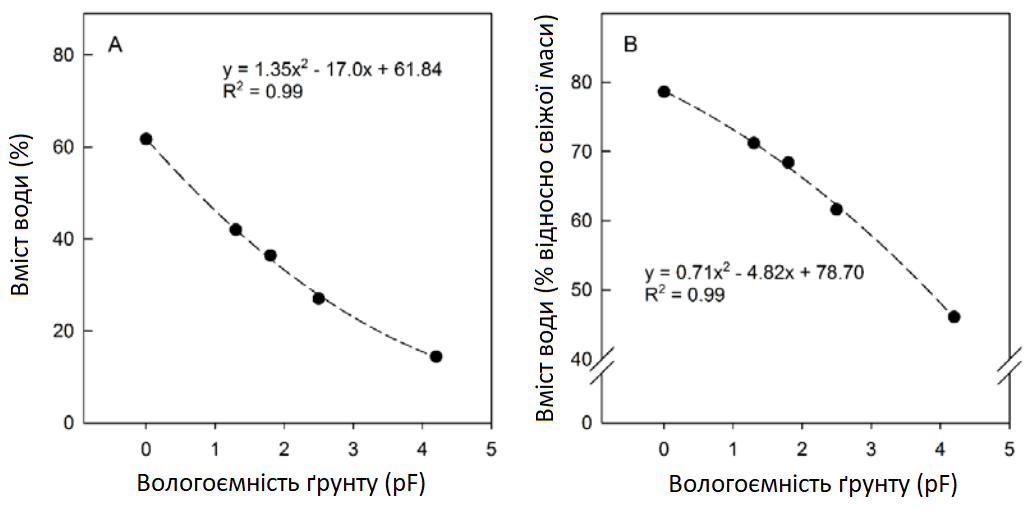
Рослини цукрового буряка поливали тричі на день по 3 хв кожну, використовуючи автоматизовану поливну систему, контрольовану за часом. Полив призвів до вмісту води приблизно 65-69% у розрахунку на свіжу масу під час росту перед експериментом, що відповідало pF (вологоємності) субстрату (log10 абсолютного значення потенціалу структури ґрунту) між 1,8 і 2,3 (Графік 1, Таблиця 2).
Таблиця 2: Характеристика субстрату Gepac Anzuchterde, тип VM
Захист рослин
Протягом усього експериментального періоду рослини залишалися вільними від шкідників та хвороб за допомогою інтегрованого захисту рослин (Таблиця 3).
Таблиця 3: Захист рослин. Огляд застосованих заходів захисту рослин протягом експериментального періоду

Дослідження розпочалося, коли рослини досягли приблизно BBCH 15-17. Потім рослини утримувались під регулярною подачею води (контролем) або зазнавали впливу посухи з подальшим відновленням зрошення (Графік 2).
Графік 2: Схематичне зображення стресу: тимчасова посуха з подальшим відновленням зрошення
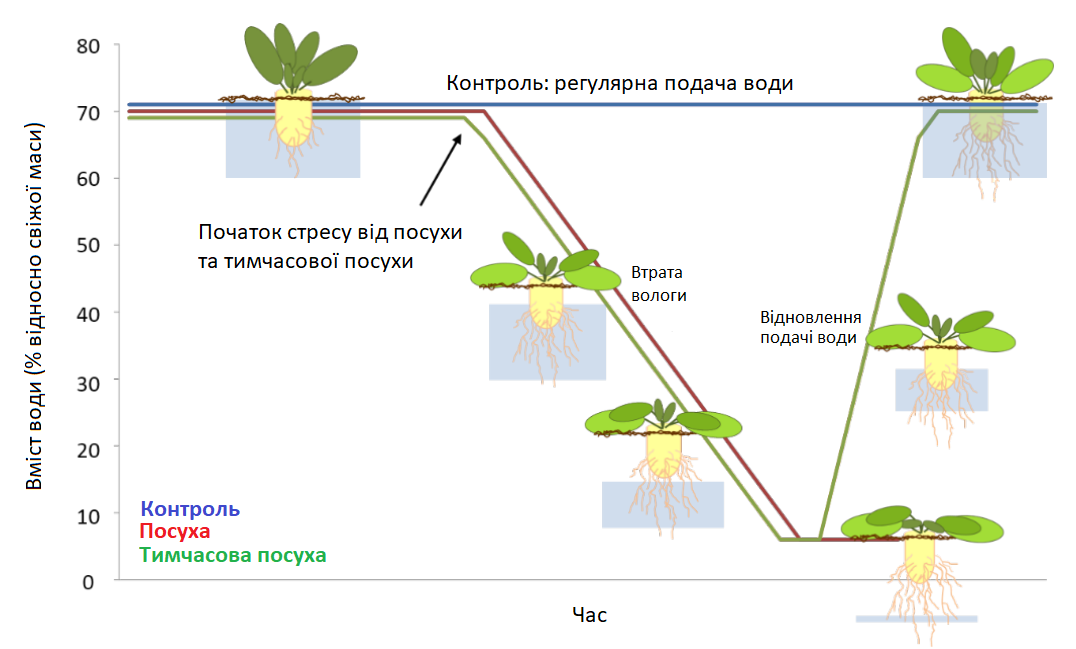
Контрольні рослини (сині) знаходилися в умовах регулярного зрошення під час усього експерименту. Стрес від посухи реалізується шляхом припинення зрошення. Рослини під дією стресу від посухи (червоні) втрачають вологу за 17 днів, тоді як рослини під дією тимчасового стресу відновлюються за 13 днів.
В усіх експериментах вміст води в ґрунті (% відносно свіжої маси) становив у середньому 65 ± 5% і підтримувався протягом усього експериментального періоду. Вміст води відповідає вологоємності субстрату 3,4, що може вважатися оптимальною подачею води (Таблиця 2).
Стрес від посухи виникав за рахунок припинення зрошення. Коренеплоди розміщувалися таким чином, що контакт із системою подачі води був неможливий. Період втрати вологи рослинами, які постраждали від посухи, становив 17 днів. Тривалість періоду відновлення рослин становила до 12 днів залежно від експерименту.
Метаболічні та фізіологічні вимірювання
Метаболічний та фізіологічний аналізи пагона проводилися на першій наймолодшій повністю розкритій парі листочків (YEL) (Графік 3). Фізіологічні вимірювання включали осмотичний потенціал (OP), відносний вміст води в листі (RWC), витік електроліту (EL), визначення малондіальдегіду (MDA), а також мікроскопічні дослідження. Кореневий аналіз проводився на найширшій частині кореня нижче крони (Графік 4). Ця частина розділялася на 2 половини. Одна половина була використана для визначення осмотичного потенціалу (OP), інша — для аналізу метаболітів на основі цільового ферменту та визначення іонів. Для аналізу метаболіту рослинний матеріал обох органів одразу заморожувався в рідкому азоті до подальшої підготовки зразків та аналізу з метою запобігання будь-якої метаболічної активності. Листовий та кореневий матеріал для метаболіту та 1H-NMR оброблялися різними способами з технічних причин. Аналіз метаболіту в пагоні проводили на свіжому (замороженому) рослинному матеріалі, тоді як аналіз 1H-NMR проводили на ліофілізованому матеріалі. Для аналізу головного кореня (обидві методики) було використано лише ліофілізований рослинний матеріал за рахунок кращої обробки.
Перед аналізом метаболіту та визначенням малондіальдегіду (MDA) листовий матеріал подрібнювався до дрібного порошку і зберігався при -80°C, тоді як для фізіологічних вимірювань (відносний вміст води в листі (RWC), витік електроліту (EL) та мікроскопія) частинки листя одразу аналізувалися або заморожувалися при температурі -20°C (OP). Для аналізу метаболіту кореневу частину різали на дрібні шматочки, потім заморожували в рідкому азоті і також зберігали при -80°С до ліофілізації. Після ліофілізації кореневий матеріал зберігали вакуумованим при -20°С до подальшого аналізу. Зберігання при -80°C та -20°C у випадку вакуумованого рослинного матеріалу є обов'язковою умовою для запобігання процесів деградації зразків.
Графік 3: Схематичне зображення пагона та кореня цукрового буряка
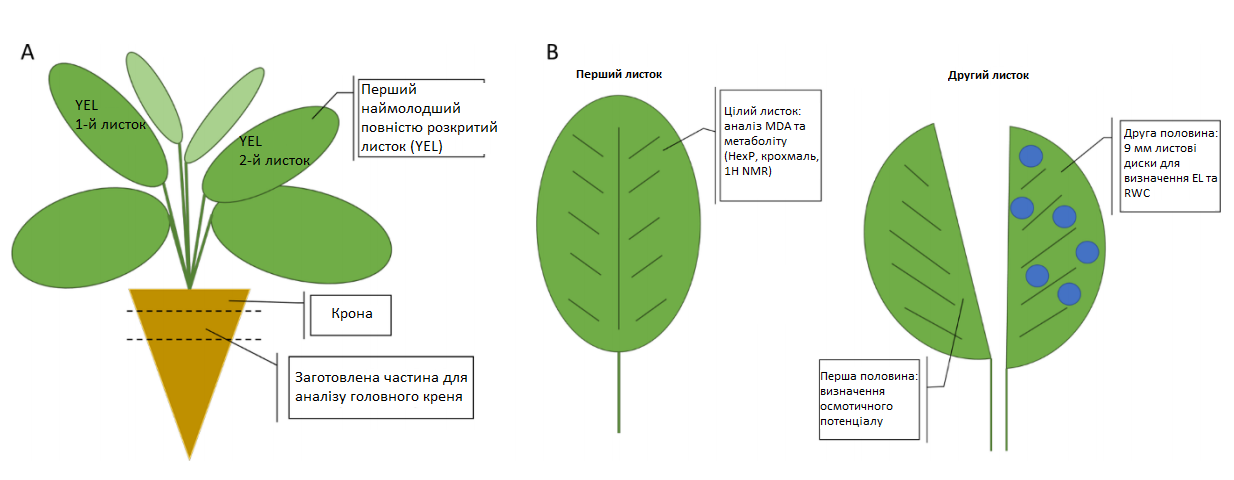
A: Огляд цукрового буряка та заготовлених частин рослини — пагона та кореня. B: Схематичне зображення YEL — наймолодшої повністю розкритої пари листочків; MDA — малодіальдегід; HexP — гексозофосфати; 1H-NMR — протонний ядерно магнітний резонанс; EL — витік електроліту; RWC — відносний вміст води.
Осмотичне регулювання в умовах посухи та відновлення зрошення
Осмотичне регулювання молодих цукрових буряків (Beta vulgaris) в умовах стресу від посухи з подальшим відновленням зрошення визначалося за аналізом метаболіту та інфрачервоною термографією.
Основна мета дослідження полягала в тому, щоб дати хронологію фізіологічних та метаболічних змін, що відбуваються в умовах посухи, та продемонструвати, як вони стосуються фенотипічного підходу (інфрачервона термографія, ІRТ). Це має забезпечити інструменти для адаптації підходів визначення фенотипу для збільшення стійкості до посухи на основі метаболічних змін. Деструктивний аналіз росту та морфології клітин, водний режим, осмотичне регулювання, метаболічні зміни та пошкодження мембрани поєднувалися з недеструктивним визначенням температури листя за допомогою інфрачервоної термографії (ІRТ) цукрових буряків, віком 6 тижнів, які піддалися впливу стресу від посухи з подальшим відновленням зрошення. Для характеристики динамічного розвитку різних фаз стресу використовувалися різні методи: хоча ІRТ дозволила виявити початкове порушення транспірації протягом 1 дня стресу, деструктивні методи дозволили розрізнити фазу метаболічної перебудови, включаючи перенаправлення потоку вуглецю на захисний механізм та наступну фазу дестабілізації мембрани та пошкодження клітин. Лише поєднання інвазивних та неінвазивних методів дозволило розмежувати повну послідовність фізіологічних змін, викликаних стресом від посухи. Це може бути особливо корисним для відбору фенотипів, стійких до посухи на ранній фазі росту. Під час відновлення зрошення пагони цукрових буряків швидко відновлювали водний режим, але пошкодження мембрани та часткове закриття продихів листя зберігалося довше, що може вплинути на можливі майбутні стресові умови. Під час початку вторинного росту кореням необхідно більше часу, щоб відновити водний режим та налагодити первинні метаболіти, ніж пагонам.
Цукровий буряк (Beta vulgaris) має досить високу стійкість до посухи, засновану на здатності зрілих рослин компенсувати тимчасову посуху повним опаданням листя та швидко відновлюватися після повернення опадів. Однак високий вихід цукру та хороша технічна якість коренеплодів вимагають достатньої кількості опадів, особливо в період розвитку вторинного кореня, а стрес від посухи є головним фактором навколишнього середовища, який відповідає за врожайність та втрати якості у виробництві цукрових буряків. В умовах навколишнього середовища зі значним випаровуванням часто спостерігається в'янення листя буряка, оскільки рослина має обмежену здатність регулювати транспірацію. Подібно до інших видів рослин, цукровий буряк накопичує осмоліти, такі як натрій (Na+), калій (К+), пролін та а-аміносполуки, такі як амінокислоти та гліцин-бетаїн, в умовах посухи. Однак високі концентрації цих осмолітів значно знижують технічну якість сировини і, таким чином, погіршують процес виробництва цукру. Із метою виявлення потенційних селекційних ознак цукрових буряків, стійких до посухи, було досліджено фізіологічні та метаболічні реакції, генетичні ресурси або генотипічні зміни сортів цукрових буряків в умовах обмеженого водопостачання. Дослідження враховували наслідки посухи під час проростання та росту сходів, або стрес від довготривалої посухи, яка впливає на всі стадії розвитку кореня, тоді як реакція молодих буряків на тимчасову посуху майже не враховувалася. Однак вирішальні фізіологічні зміни, такі як початок росту вторинного кореня, відбуваються на стадії молодих рослин 14-18 BBCH. Після цього починається накопичення сахарози, що важливо для кінцевого виходу цукру. Короткі тимчасові посушливі періоди можуть негативно позначитися на цій ранній, але досить важливій, стадії розвитку, знизити кінцеву врожайність і, отже, економічний прибуток. У Центральній Європі, на відміну від багатьох посушливих країн нижчих широт, посушливі періоди часто є не періодичними, але можуть виникати на будь-якій стадії розвитку цукрових буряків. Вони часто перериваються випадковими опадами, які варіюються від поодиноких, як правило, сильних злив, до декількох днів або тижнів та є нерівномірними. За прогнозами глобального потепління, в Європі очікується збільшення кількості екстремальних погодних явищ, зміна сезонних та регіональних опадів, а саме скорочення літніх опадів та супутнє збільшення числа тимчасових посушливих періодів на початку вегетаційного розвитку рослин. У районах із періодичними періодами посухи врожайність буряків сильно залежить від здатності рослин відновлюватися після поновлення зрошення, що включає механізми відновлення росту пагонів і коренів та осмотичного гомеостазу, відновлення тканин, пошкоджених окислювальним стресом, та налаштування метаболізму. Неповне або повільне відновлення може змінити реакцію рослини на наступні стресові умови — явище, яке ми спостерігали в експериментах із цукровими буряками, що зазнавали повторних циклів посухи лише з короткими періодами відновлення. Крім того, в польовому експерименті спостерігалося неповне відновлення осмотичного пристосування цукрових буряків після довготривалої посухи з подальшим відновленням зрошення і вважалося критичним для технічної якості цукру. Селекційні програми для схильних до посухи умов навколишнього середовища потребують раннього та неінвазивного виявлення стресу для полегшення характеристики сортів у визначенні фенотипу та в польових випробуваннях. Швидке виявлення стресу необхідне для ефективного та точного налаштування зрошення. Було розроблено велику кількість неінвазивних сенсорних методик для виявлення стесу у високопропускних фенотипувальних установках. Інфрачервона термографія (ІRТ) може бути використана для визначення водного режиму рослин, реакцій на сольовий стрес або появу інфекційних збудників на ранніх стадіях розвитку рослин. Досі існує розрив між зовнішнім фенотипом та відповідними фізіологічними та метаболічними процесами, які, однак, являють собою "внутрішній фенотип" та інтегрують сигнали навколишнього середовища у відповідь клітин. Отже метою дослідження було поєднання методів недеструктивної та деструктивної діяльності як крок до встановлення зв’язку між неінвазивною візуалізацією та динамікою фізіологічних та обмінних процесів. При такому методологічному підході дослідження стосувалося двох цілей: (І) визначення динаміки фізіологічних та метаболічних реакцій на стрес від тривалої посухи на рослинах Beta vulgaris на ранніх етапах росту вторинного кореня та (ІІ) дослідження динаміки відновлення різних фізіологічних процесів після відновлення зрошення.
Результати
Ріст та розвиток
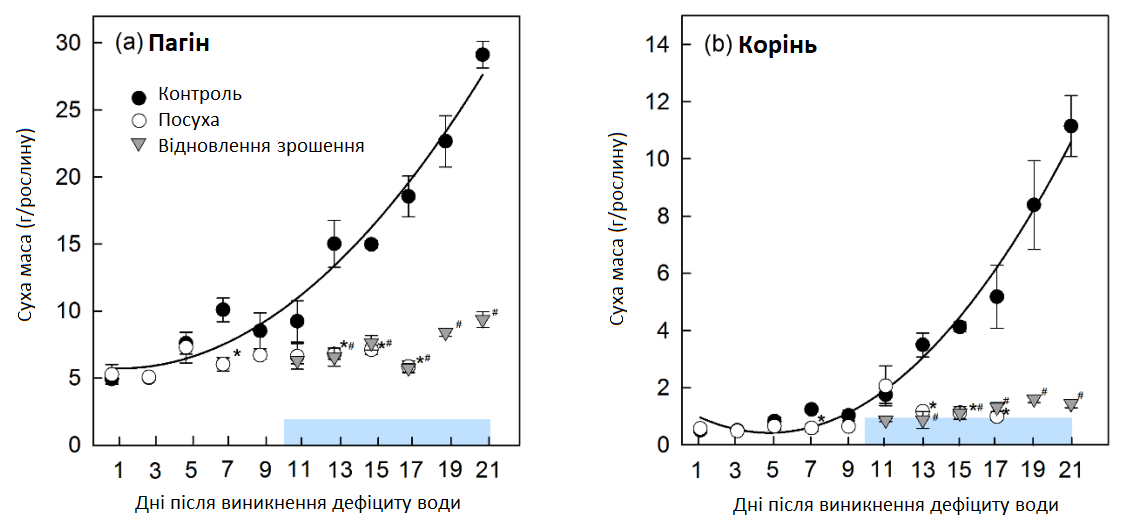
В умовах контролю ріст коренів і пагонів збільшувався в геометричній прогресії протягом усього експерименту (Графік 2), рослина розвинулася з рівня BBCH 16-17 в 1-й день до BBCH 19 на 21-й день (Графік 3). В умовах стресу ріст рослин сильно гальмувався, починаючи з 5-7 дня (пагони) та 11-13 дня (корінь) після припинення зрошення (Графік 2). В'янення старого листя стало помітним через 5 днів і сильно позначилося на всіх повністю розкритих листках через 9 днів (Графік 3). Однак наймолодші листочки залишалися зеленими та в стані хорошого тургору до 11 дня. Наприкінці посухи (13-17 день) старе листя стало хлоротичним та повністю висохло. Ріст коренів та пагонів відновився через 5-7 днів після відновлення зрошення (Графік 2). Наймолодші листочки відновили тургор протягом одного дня, а в'янення старішого листя зникло протягом 7 днів (Графік 3).
Графік 2: Суха маса (DW) молодих пагонів цукрових буряків та коренеплодів. Суха маса (DW) (а) пагонів молодих цукрових буряків та (b) коренів в умовах регулярного зрошення (контроль, темні кола), стресу від тривалої посухи (світлі кола) та відновлення зрошення (трикутники).
Температура листя
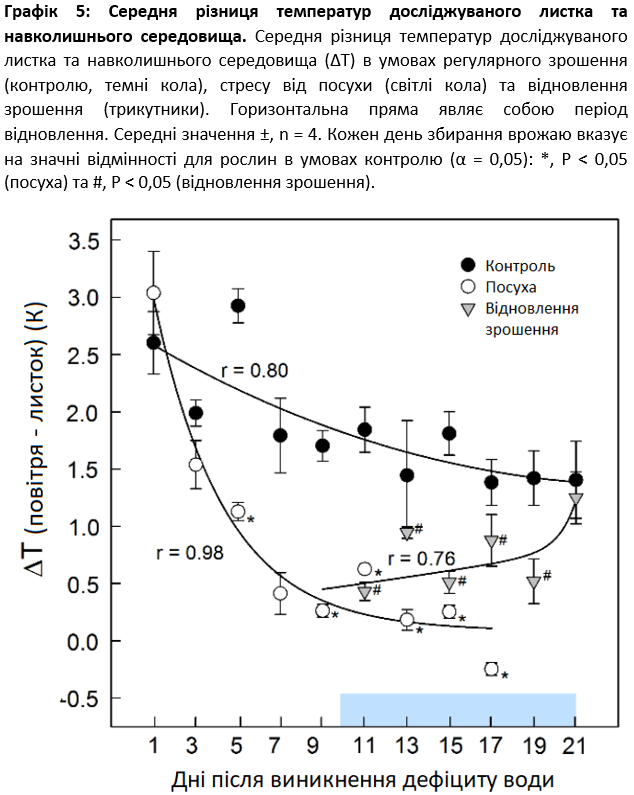
В умовах контролю рослини мали відносно постійну температуру листя (за винятком біологічно найстарішого листя) протягом усього експерименту, тоді як в умовах посухи температура листя постійно зростала і досягла температури навколишнього середовища наприкінці стресового періоду (Графік 4), коли рослини були візуально повністю висушені (Графік 3). Для спостереження за зміною температури листя в часі використовувалася температурна різниця (∆T) між середньою температурою наймолодшої повністю розкритої пари листочків (YEL) та температурою повітря (Графік 5). Щодо контрольованих рослин, ∆T коливалася від 2,9 до 1,4 К протягом всього експерименту, але зменшилася з 3,0 до -0,3 К в умовах посухи. Через 5 днів після припинення зрошення різниця була значною (P ≤ 0,024) (Графік 5). Основне падіння ∆T відбулося протягом перших 3 днів посухи із середньою швидкістю 0,50 К в день-1 і тривало при середніх швидкостях 0,18 К/добу до 7 дня та 0,06 К/добу-1 до 17 дня відповідно. Під час відновлення зрошення ∆T почала зростати після періоду відставання у 2 дні, а контрольні значення були досягнуті через 12 днів.
Графік 3: RGB-зображення цукрових буряків протягом експериментального періоду. RGB-зображення молодих рослин цукрових буряків в умовах регулярного зрошення (контролю), стресу від посухи та відновлення зрошення, день 1, 5, 9, 11, 17 та 21. Стрілка вказує на досліджуваний листок.

Графік 4: Інфрачервоні теплові зображення. Інфрачервоні теплові зображення молодих цукрових буряків в умовах регулярного зрошення (контролю), посухи та відновлення зрошення, день 1, 5, 9, 11, 17 та 21. Стрілка вказує на досліджуваний листок.

 Водний режим рослин та стабільність мембрани
Водний режим рослин та стабільність мембрани
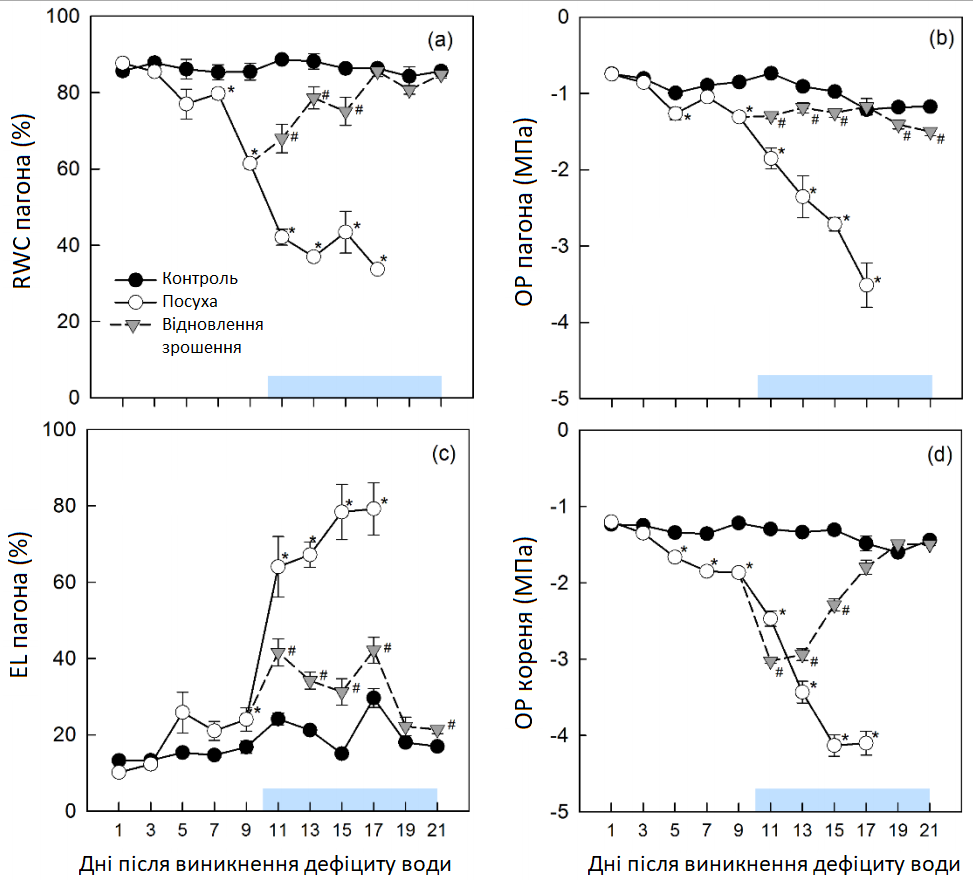
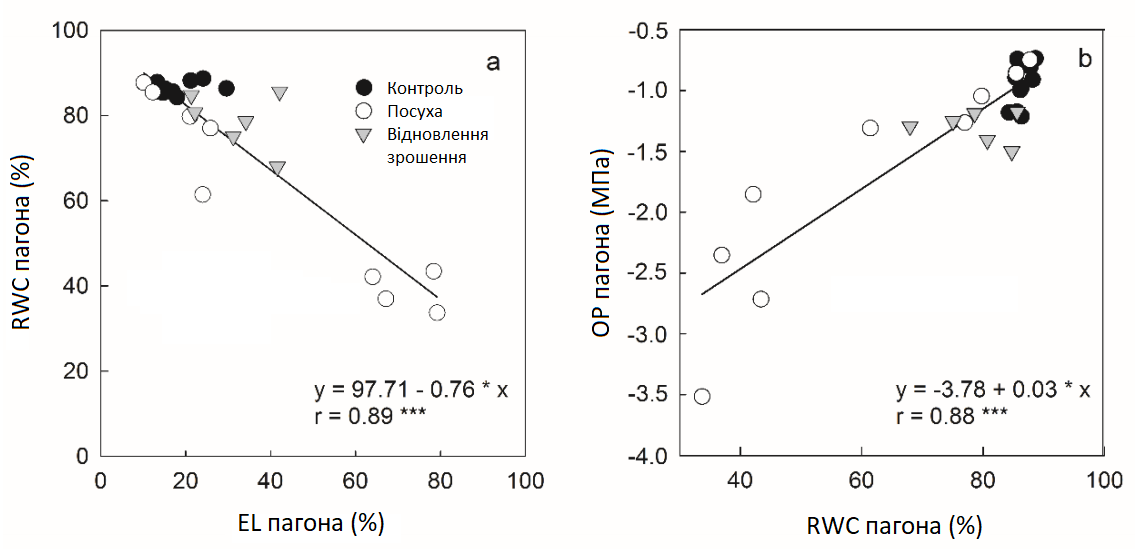
В умовах контролю відносний вміст води пагона (RWCS) та осмотичний потенціал пагона (OPS) та кореня (OPR) були стабільними із середніми значеннями 86 ± 1%, -1,0 ± 0,2 МПа та -1,4 ± 0,1 МПа відповідно (Графік 6). Усі ці параметри відповідали умовам посухи із аналогічною часовою динамікою. Протягом перших 3 днів посухи вони не змінювалися, потім повільно зменшувалися до 7 чи 9 дня, а потім значно знижувались (Графік 6). Між 7 та 11 днями нахили відповідних кривих істотно змінилися. RWCS досягнув 42 ± 4% через 11 днів і залишався на цьому рівні до кінця експерименту, тоді як через 17 днів кінцевий ОP становив -3,5 ± 0,5 МПа в пагонах і -4,1 ± 0,3 МПа в коренях. Криві реакцій відображаються реакцією витоку електроліту пагона (ELS) на посуху з помірним, але значним збільшенням з 3 по 9 день та набагато більшим збільшенням з 9 по 17 день, що свідчить про сильне пошкодження мембрани під час цієї фази стресу (Графік 6). Загалом високо значущі лінійні кореляції спостерігалися між RWCS та ELS (r = 0,89), а також OPS та RWCS (r = 0,88, Графік 7). Пагони та корені по-різному реагували на відновлення зрошення. Оскільки спричинене посухою зниження RWCS та OPS негайно припинилося, а збільшення ELS між 9 та 11 днями зменшилось на 53%, OPR продовжував знижуватися ще 2 дні до початку відновлення (Графік 6). Відносний вміст води (RWC) та осмотичний потенціал (OP) залишалися нижчими контрольних рівнів принаймні 6-8 днів, тоді як ELS повністю не повернувся до контрольних значень до кінця періоду відновлення.
Графік 6: RWC, OP та EL пагона та OP кореня. (a) відносний вміст води (RWC), (b) осмотичний потенціал (OP), (c) витік електроліту (EL) молодого цукрового буряка та (d) осмотичний потенціал молодого кореня в умовах зрошення (контролю, темні кола), стресу від посухи (світлі кола) та відновлення зрошення (трикутники). Горизонтальна пряма являє собою період відновлення. Середні значення ±, n = 4. Кожен день збирання врожаю вказує на значні відмінності для рослин в умовах контролю (α = 0,05): *, P < 0,05 (посуха) та #, P < 0,05 (відновлення зрошення).
Графік 7: Кореляційний аналіз відносного вмісту води, витоку електроліту та осмотичного потенціалу. Кореляція між витоком електроліту (EL) та відносним вмістом води (RWC) (A), між RWC та осмотичним потенціалом (OP) (B) пагонів цукрових буряків в умовах зрошення (контролю, темні кола), стресу від посухи (світлі кола) та відновлення зрошення (трикутники). Середні значення ±, n = 4.
Зв'язок між температурою та водним режимом листя
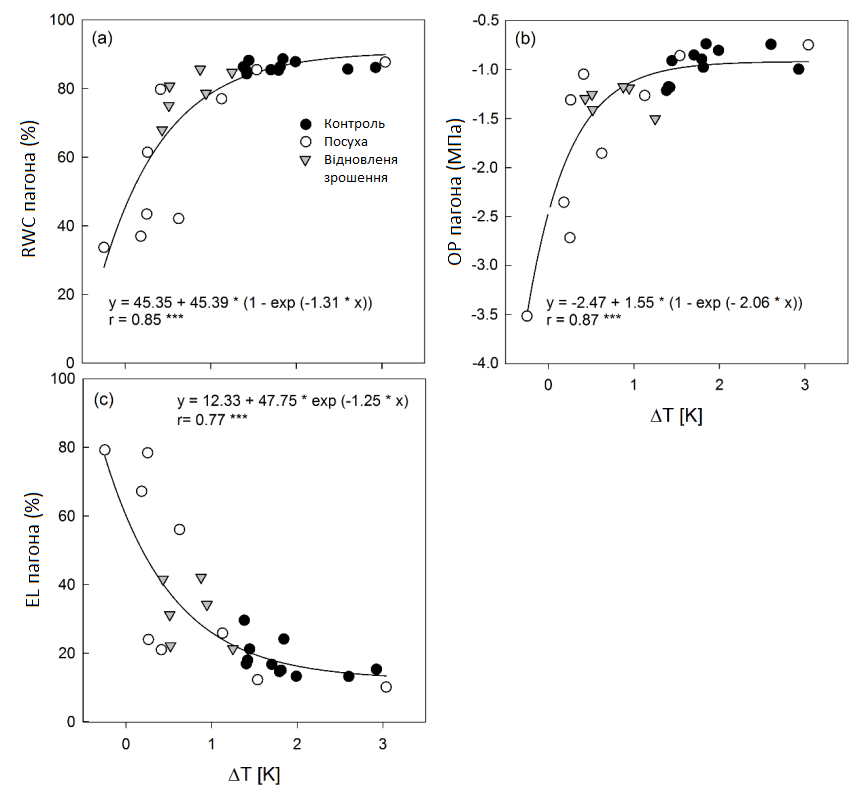
Для визначення природи взаємозв'язку між ΔT та водним режимом листя (RWCS, OPS) або пошкодженням мембрани (ELS) були співвіднесені середні значення кожного параметра. У всіх випадках були отримані високо значущі (P < 0,001) експоненціальні співвідношення (Графік 8).
Графік 8: Кореляційний аналіз середньої різниці температур (ΔT) та RWC, OP та EL. Кореляційний аналіз середньої різниці температури досліджуваного листка та навколишнього середовища (∆T) та (a) відносного вмісту води (RWC), (b) осмотичного потенціалу (OP) та (c) витоку електроліту (EL) пагонів цукрових буряків в умовах зрошення (контролю, темні кола), стресу від посухи (світлі кола) та відновлення зрошення (трикутники). Середні значення ±, n = 4.